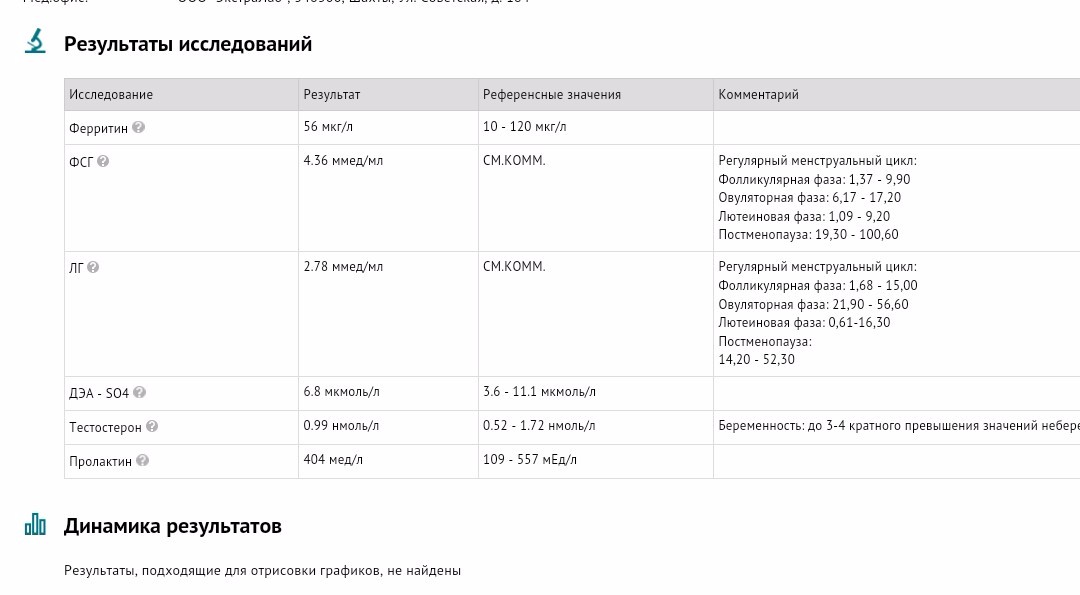
Дегидроэпиандростерон-сульфат (ДЭА-S04, ДЭА-С, Dehydroepiandrosterone sulfate, DHEA-S)
Что такое дегидроэпиандростерон-сульфат (ДЭА-S04, ДЭА-С, Dehydroepiandrosterone sulfate, DHEA-S)?
Дегидроэпиандростерон-сульфат (ДЭА-S04, ДЭА-С, Dehydroepiandrosterone sulfate, DHEA-S) – это андроген, то есть мужской половой гормон, который вырабатывается в основном надпочечниками (парными эндокринными органами, расположенными на верхнем полюсе почек). Дегидроэпиандростерон-сульфат присутствует в организме как мужчин, так и женщин, небольшая часть этого гормона вырабатывается яичками и яичниками. Этот гормон может превращаться в организме в более сильные андрогены, например, тестостерон, и в эстроген (женский половой гормон).
Дегидроэпиандростерон-сульфат играет роль в развитии вторичных половых признаков у мальчиков в период полового созревания, таких как оволосение по мужскому типу, изменение тембра голоса и т.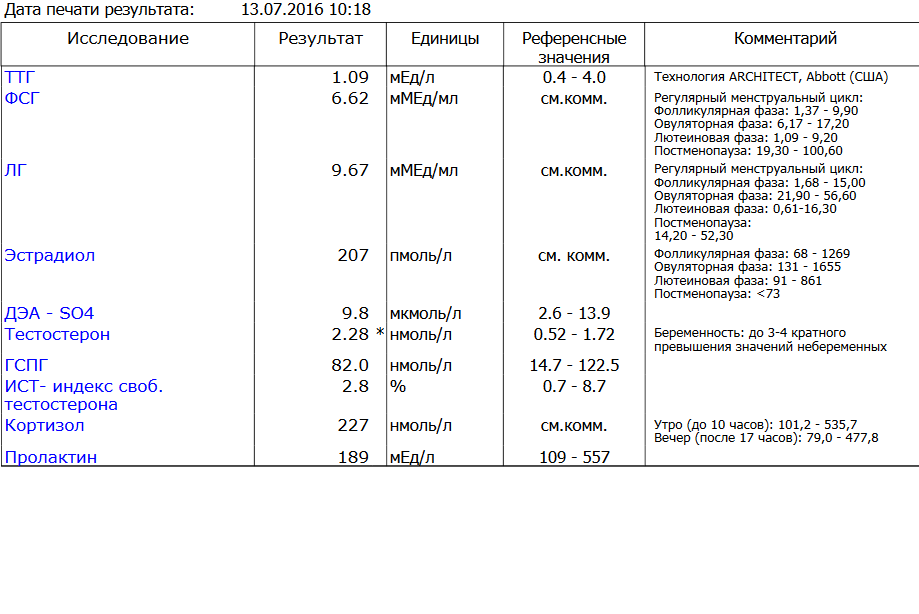 д.
д.
Для чего определяют уровень дегидроэпиандростерон-сульфата в крови?
Определение уровня дегидроэпиандростерон-сульфата используют для оценки функции надпочечников (к повышению показателя приводят опухоли надпочечников и гиперплазия – увеличение количества клеток; снижение показателя свидетельствует о недостаточной функции надпочечников в отношении выработки половых гормонов), реже – для выявления опухоли яичников, которая также может продуцировать слишком большое количество этого гормона. В этом случае симптомы патологии будут включать в себя избыточный рост волос на лице и теле (гирсутизм), развитие мужских черт (вирилизация, или маскулинизация) и нарушения цикла вплоть до отсутствия менструации (аменореи) и бесплодия.
Как правило, для наиболее точной диагностики тест на концентрацию дегидроэпиандростерон-сульфата выполняют вместе с анализами на уровень половых гормонов (тестостерона, эстрогена).
При каких заболеваниях изменяется уровень дегидроэпиандростерон-сульфата в крови?
Уровень дегидроэпиандростерон-сульфата повышается при гиперплазии или опухоли надпочечников (как доброкачественной, так злокачественной), синдроме поликистозных яичников (СПКЯ – гормональное заболевание женщин детородного возраста, при котором в яичниках появляются множественные полости, заполненные жидкостью – кисты, а симптомы включают нарушение менструального цикла вплоть до аменореи, гирсутизм, склонность к полноте и др.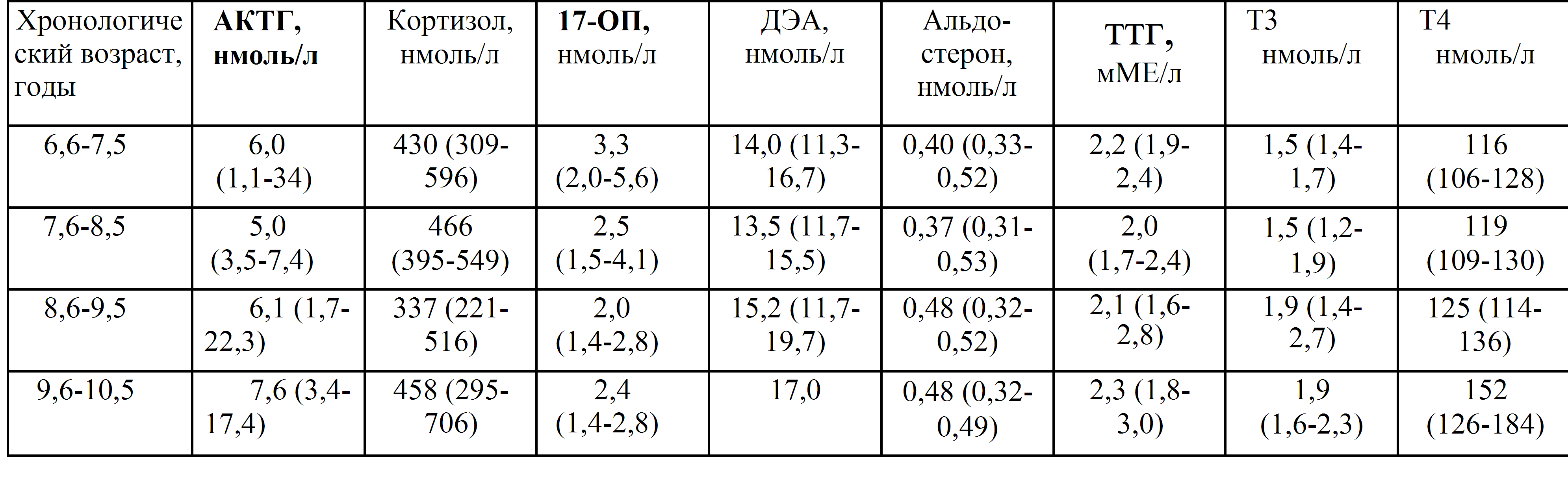 ), при болезни Иценко – Кушинга и эктопических (расположенных вне гипофиза) опухолях, продуцирующих АКТГ (адренокортикотропный гормон).
), при болезни Иценко – Кушинга и эктопических (расположенных вне гипофиза) опухолях, продуцирующих АКТГ (адренокортикотропный гормон).
Уровень дегидроэпиандростерон-сульфата снижается при дисфункции и заболеваниях надпочечников (болезни Аддисона, синдроме Иценко – Кушинга и др.), гипопитуитаризме (когда гипофиз не вырабатывает некоторые или все гормоны).
Почему результат анализа может быть некорректным?
На концентрацию дегидроэпиандростерон-сульфата в крови может влиять прием дегидроэпиандростерон-сульфатных добавок или некоторых препаратов, например, содержащих гестагены (аналоги полового гормона прогестерона). О принимаемых постоянно или периодически медикаментах следует сообщить врачу, назначившему исследование.
Интерпретация результатов исследований содержит информацию для лечащего врача и не является диагнозом. Информацию из этого раздела нельзя использовать для самодиагностики и самолечения. Точный диагноз ставит врач, используя как результаты данного обследования, так и нужную информацию из других источников: анамнеза, результатов других обследований и т.д.
Трактовка результатов определения уровня Дегидроэпиандростерон-сульфата в сыворотке крови
Единицы измерения: мкмоль/л.
Альтернативные единицы: мкг/дл.
Переcчет единиц: мкг/дл х 0,02714 ==> мкмоль/л.
Референсные значения
| Возраст | Пол | ДЭА-SO4, мкмоль/л |
| 1-5 лет | оба | <2,48 |
| 5-10 лет | оба | <4,86 |
| 10-14 лет | оба | 0,7-10,32 |
| 14-20 лет | жен | 1,7-13,4 |
| 1,2-10,4 | ||
| 20-25 лет | жен | 3,6-11,1 |
| муж | 6,5-14,6 | |
| 25-35 лет | жен | 2,6-13,9 |
| муж | 4,6-16,1 | |
| 35-45 лет | жен | 2,0-11,1 |
| муж | 3,8-13,1 | |
| 45-55 лет | жен | 1,5-7,7 |
| муж | 3,7-12,1 | |
| >55 лет | жен | 0,8-4,9 |
| муж | 1,3-9,8 |
Повышение значений
- Адреногенитальный синдром.

- Опухоли коры надпочечников.
- Эктопические АКТГ-продуцирующие опухоли.
- Болезнь Кушинга (гипоталамо-гипофизарный генез).
- Фетоплацентарная недостаточность.
- Гирсутизм женщин (при нормальном показателе заболевание не связано с патологией надпочечников).
- Угроза внутриутробной гибели плода.

Понижение значений
- Гипоплазия надпочечников плода (концентрация в крови беременной женщины).
- Внутриутробная инфекция.
- Приём таких препаратов как гестагены.
Анализ на ДГЭА — SO4
ДГЭА-С – основной стероид, секретируемый корой надпочечников и являющийся предшественником биосинтеза тестостерона, андростендиона и эстрогенов (эстрона и эстрадиола). ДГЭА -C обладает относительно слабой андрогенной активностью, составляющей примерно 10% от уровня тестостерона. Однако, его биологическая активность усиливается благодаря относительно высоким концентрациям в сыворотке – в 100 или 1000 раз превосходящим тестостерон, а также из-за слабой афинности к глобулину, связывающему половые гормоны (SHBG). Содержание ДГЭА-С в сыворотке является маркером синтеза андрогенов надпочечниками.
ДГЭА -C обладает относительно слабой андрогенной активностью, составляющей примерно 10% от уровня тестостерона. Однако, его биологическая активность усиливается благодаря относительно высоким концентрациям в сыворотке – в 100 или 1000 раз превосходящим тестостерон, а также из-за слабой афинности к глобулину, связывающему половые гормоны (SHBG). Содержание ДГЭА-С в сыворотке является маркером синтеза андрогенов надпочечниками.
Поскольку лишь незначительная часть гормона образуется половыми железами, измерение ДГЭА-С может помочь в определении локализации источника андрогенов. Если у женщин наблюдается повышенный уровень тестостерона, то с помощью определения ДГЭА-С можно установить, связано ли это с болезнью надпочечников или с заболеванием яичников.
Уровень ДГЭА-С не зависит от фазы менструального цикла, не имеет циркадного ритма секреции. Значения у мужчин и женщин с возрастом прогрессивно снижаются.
Во время беременности ДГЭА-C вырабатывается корой надпочечников матери и плода и служит предшественником для синтеза эстрогенов плаценты.
Определение ДГЭА-C заменяет определение 17-КС в моче при оценке выработки надпочечниками андрогенов.
Показания к назначению:
- Дифференциальная диагностика заболеваний надпочечников и заболеваний яичников с повышенным уровнем тестостерона в крови
- Адреногенитальный синдром
- Опухоли коры надпочечников
- Эктопические АКТГ-продуцирующие опухоли
Когда уровень ДГЭА-С повышен:
- Адреногенитальный синдром
- Опухоли коры надпочечников
- Эктопические АКТГ-продуцирующие опухоли
- Болезнь Кушинга
- Синдром поликистозных яичников
- Гирсутизм у женщин
Когда уровень ДГЭА-С понижен:
- Недостаточность надпочечников
- Задержка полового созревания
- Прием препаратов: глюкокортикоидов, пероральных контрацептивов
Необходимо воздержаться от приема пищи в течение 2-3 часов. При сохраненном м/цикле — на 5-7 д. м/ц, если другие сроки не указаны врачом
При сохраненном м/цикле — на 5-7 д. м/ц, если другие сроки не указаны врачом
Дегидроэпиандростерон-сульфат (ДЭА-S04, ДЭА-С, Dehydroepiandrosterone sulfate, DHEA-S)
Исследуемый материал Сыворотка крови
Метод определения Твёрдофазный хемилюминесцентный иммуноанализ
Данное исследование можно выполнить в режиме «Приоритет» — результаты до 14 часов в медицинских офисах по следующим адресам:- г. Алматы, 5 мкрн д 17 (Абая/Алтынсарина).
- г. Алматы, ул. Мендикулова (мкр. Самал-2), д.3
- г. Алматы, Бостандыкский р-н, мкр. Орбита — 4, д. 11
- г. Алматы, пр-кт Абая, 89
 Алматы, 5 мкрн д 17 (Абая/Алтынсарина).
Алматы, 5 мкрн д 17 (Абая/Алтынсарина). Андрогенный гормон надпочечников.
Вырабатывается в коре надпочечников. Уровень этого гормона является адекватным показателем андроген-синтетической активности надпочечников. Гормон обладает лишь слабым андрогенным действием, однако, в процессе его метаболизма в периферических тканях образуются тестостерон и дигидротестостерон. Не обнаруживает заметных суточных колебаний и имеет низкую скорость клиренса.
Во время беременности вырабатывается корой надпочечников матери и плода и служит предшественником для синтеза эстрогенов плаценты. Его уровень повышается к периоду полового созревания, а затем плавно снижается по мере выхода человека из репродуктивного возраста. Во время беременности уровень этого гормона также снижается.
Во время беременности уровень этого гормона также снижается.
Определение ДЭА-SO4 заменяет определение 17-КС в моче при оценке выработки надпочечниками андрогенов. В яичниках синтеза ДЭА-сульфата не происходит (поэтому тест применяется для определения источника гиперандрогенемии в организме женщины).
Состояние репродуктивной системы и мониторинг беременности
Дегидроэпиандростерон-сульфат (ДЭА-S04, ДЭА-С, Dehydroepiandrosterone sulfate, DHEA-S)
Описание
Дегидроэпиандростерон-сульфат (ДЭА-SO4) – это андроген, мужской половой гормон, который присутствует в крови как у мужчин, так и у женщин. Он играет важную роль в развитии вторичных мужских половых признаков при половом созревании и может преобразовываться в тестостерон и эстрадиол. ДЭА-SO4 вырабатывается корой надпочечников, его производство контролируется адренокортикотропным гормоном (АКТГ).
Единицы измерения
Мкг/дл (микрограмм на децилитр)
Подготовка к исследованию
- Не принимать пищу в течение 2-3 часов перед исследованием, можно пить чистую негазированную воду.
- Прекратить прием стероидных и тиреоидных гормонов за 48 часов до исследования (по согласованию с врачом).
- Исключить физическое и эмоциональное перенапряжение за 24 часа до сдачи крови.
- Не курить в течение 3 часов до исследования.
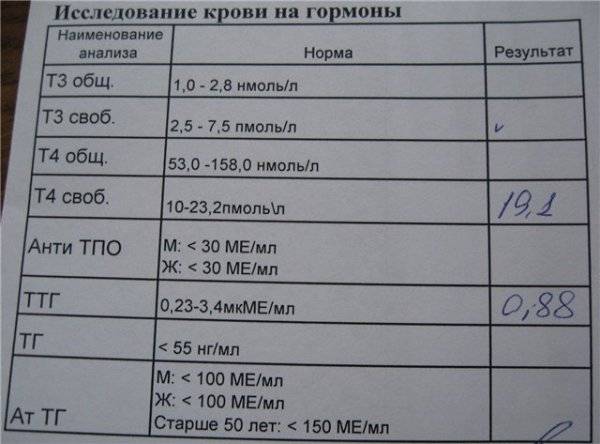
Нормальные значения (референсы)
Возраст | Референсные значения |
Дети | |
< 1 недели | 108 — 607 мкг/дл |
1-4 недели | 31,6 — 431 мкг/дл |
1-12 месяцев | 3,4 — 124 мкг/дл |
1-5 лет | 0,47 — 19,4 мкг/дл |
5-10 лет | 2,8 — 85,2 мкг/дл |
Женщины | |
10-15 лет | 33,9 — 280 мкг/дл |
15-20 лет | 65,1 — 368 мкг/дл |
20-25 лет | 148 — 407 мкг/дл |
25-35 лет | 98,8 — 340 мкг/дл |
35-45 лет | 60,9 — 337 мкг/дл |
45-55 лет | 35,4 — 256 мкг/дл |
55-65 лет | 18,9 — 205 мкг/дл |
65-75 лет | 9,4 — 246 мкг/дл |
> 75 лет | 12 — 154 мкг/дл |
Мужчины | |
10-15 лет | 24,4 — 247 мкг/дл |
15-20 лет | 70,2 — 492 мкг/дл |
20-25 лет | 211 — 492 мкг/дл |
25-35 лет | 160 — 449 мкг/дл |
35-45 лет | 88,9 — 427 мкг/дл |
45-55 лет | 44,3 — 331 мкг/дл |
55-65 лет | 51,7 — 295 мкг/дл |
65-75 лет | 33,6 — 249 мкг/дл |
> 75 лет | 16,2 — 123 мкг/дл |
Приемлемые уровни ДЭА-SO4 и других андрогенов могут свидетельствовать о нормальном функционировании надпочечников. Гораздо реже количество ДЭА-SO4 бывает в норме при опухоли надпочечника, но при этом не происходит секретирования гормона. При синдроме поликистозных яичников уровень ДЭА-SO4 может быть как повышенным, так и нормальным, поскольку это заболевание обычно связано с производством андрогенов яичниками (главным образом тестостерона).
Гораздо реже количество ДЭА-SO4 бывает в норме при опухоли надпочечника, но при этом не происходит секретирования гормона. При синдроме поликистозных яичников уровень ДЭА-SO4 может быть как повышенным, так и нормальным, поскольку это заболевание обычно связано с производством андрогенов яичниками (главным образом тестостерона).
Повышенный уровень ДЭА-SO4 может свидетельствовать об опухоли коры надпочечников, раке либо гиперплазии надпочечников. Он не является диагностическим признаком какой-то определенной болезни, однако указывает на необходимость дальнейшего обследования для выявления причин гормонального дисбаланса.
Низкий уровень ДЭА-SO4 бывает вызван дисфункцией надпочечников или гипопитуитаризмом, нарушениями, являющимися причиной понижения уровня питуитарных гормонов, которые, в свою очередь, регулируют производство и секрецию гормонов, производимых надпочечниками.
Дегидроэпиандростерон-сульфат (ДЭА-S04, Dehydroepiandrosterone sulfate, DHEA-S)
ФИО*:
Телефон*:
Необходимая услуга или врач: Комиссия для иностранных гражданМедицинская комиссия для трудоустройстваПриём аллергологаПриём врача УЗД (УЗИ)Приём гастроэнтерологаПриём гинекологаПриём дерматологаПриём детского аллергологаПриём детского гастроэнтерологаПриём детского гинекологаПриём детского кардиологаПриём детского психиатраПриём детского психологаПриём детского пульмонологаПрием детского стоматологаПриём детского урологаПриём детского хирургаПриём кардиологаПриём косметологаПриём логопедаПриём массажистаПриём неврологаПриём онкологаПриём ортодонтаПриём ортопедаПриём отоларингологаПриём офтальмологаПрием педиатраПриём проктологаПриём профпатологаПриём психиатра-наркологаПрием психотерапевтаПриём пульмонологаПриём ревматологаПриём сосудистого хирургаПриём стоматологаПриём терапевтаПриём трихологаПриём урологаПриём физиотерапевтаПриём хирургаПриём эндонкринологаСкрининг гинекологаЭКГ-ХолтерАбдулов Игорь АнатольевичАбдулова Валентина ИвановнаАкимова Нина ВикторовнаАксенов Кирилл СергеевичАлексеенко Мария НиколаевнаАннаев Максат ГеокчаевичБалашов Александр ВячеславовичБатова Елена ВикторовнаБелкина Анжелика СтаниславовнаБеляков Алексей СергеевичБойкова Мария ОлеговнаБондаренко Марина ВалерьевнаБосых Владимир ГеоргиевичБрага Раиса ИвановнаБулатов Дмитрий АлександровичБурбот Любовь ВикторовнаВдовина Елена ВитальевнаВиноградова Оксана НиколаевнаВласова Светлана АлександровнаГауст Анисья РадифовнаГвозденко Сергей ФедоровичГорбачев Илья СергеевичГригорьев Сергей АлексеевичГригорьева Анна БорисовнаДавыдова Надежда ВасильевнаДавыдова Елена ЮрьевнаДевяткина Варвара ПавловнаДеменкова Виктория ВладимировнаДинамарка Карина ФернандовнаДобко Зоя ГригорьевнаДустаметова Сабина ДустаметовнаЕгоренко Елена АнатольевнаЕжова Любовь ГеннадьевнаЖуков Семен АндреевичЗамостян Анна ДмитриевнаЗейналов Эльмар Кафар оглыЗинченко Светлана ИвановнаЗмановская Татьяна ЛеонидовнаИванов Александр АлександровичИвашкина Екатерина ДмитриевнаИкромов Сухробжон НасруллоевичИркова Ирина АнатольевнаКалашникова Елена ПетровнаКалинина Анна СергеевнаКандрашкина Екатерина ЕвгеньевнаКиселев Игорь ЕвгеньевичКиселева Наталия СтаниславовнаКозлова Инна ИвановнаКокорина Оксана ВалериевнаКолодина Юлия МихайловнаКольдин Алексей ВладимировичКорнев Алексей ВячеславовичКорчагина Антонина НиколаевнаКострюкова Лариса НиколаевнаКравцова Марина ЮрьевнаКрасулина Ольга АлександровнаКрюкова Оксана АндреевнаКудеева Оксана ВикторовнаКузьмина Елена НиколаевнаКулагина Татьяна СтаниславовнаКутлахметов Айрат АзгаровичЛихачев Никита ЕвгеньевичЛогинов Виталий АлександровичЛукьянова Екатерина ЮрьевнаМакаркин Владимир СергеевичМальцев Максим ЮрьевичМарченко Лилия ВладимировнаМатях Игорь ИгоревичМаханов Рустам ХамиджоновичМаханова Ольга БазаровнаМережко Вероника ИгоревнаМерзова Фируза РафиковнаМещеряков Михаил ВикторовичМиронова Марина АнатольевнаМихайлов Дмитрий ВладимировичМолчанова Надежда ПетровнаНазарчук Светлана НиколаевнаНайман Сергей ПавловичНикулин Павел НиколаевичНовиков Олег ЛеонидовичНовиков Алексей НиколаевичНовикова Ирина ВладимировнаНовикова Елена ВячеславовнаОдинец Лидия ФедоровнаОрлова Ольга АлександровнаОхотина Инна ИгоревнаПавлов Владимир СергеевичПанченко Ирина АнатольевнаПапин Александр ГеоргиевичПоздняков Евгений ГеннадьевичПопов Сергей ВикторовичПоспелова Рита АнатольевнаПучкова Наталья АлександровнаРепин Павел НиколаевичРешетникова Татьяна ПетровнаРогожин Павел СергеевичРогожина Екатерина ГеннадьевнаРостиков Олег ВячеславовичРудаева Любовь МихайловнаРыкова Марина ВладимировнаСкорнякова Ирина ИгоревнаСмирнова Людмила АлександровнаСоколова Татьяна ФедоровнаСорокина Елена КонстантиновнаСтепанова Виктория СергеевнаСтепашкина Анастасия СергеевнаСтроганова Тамара ИвановнаСычева Полина АлександровнаТарарышкин Дмитрий АлександровичТарахтиева Наталья ВасильевнаТерехина Наталья ВладимировнаТестовый Врач КлиникиТетерина Елена ВалерьевнаТихонов Алексей ВладимировичТутунина Елена ВладимировнаФедосеева Надежда ВикторовнаФокина Алена АлексеевнаХарламов Павел ВикторовичЧапчикова Ольга АлександровнаЧеремина Виктория ВикторовнаЧернецкая Инесса ИвановнаЧернова Любовь ВладимировнаЧижов Михаил СергеевичЧичерина Валентина ВикторовнаШаповалова Нина БорисовнаШкурлатов Сергей НиколаевичШтейн Юлия СергеевнаЩекочихина Тамара ВикторовнаЩекочихина Тамара ВикторовнаЩербак Валерия НиколаевнаЯгодина Екатерина Антоновна
Клиника Все клиникиДетская поликлиника «Живица+»Многопрофильная клиника «Живица+»Медицинский центр «Живица+» в ГолутвинеМедицинский центр «Живица+» на Окском
Предпочтительное время приема:
Детский врач Принимает детей
Отправляя заявку, вы даете согласие на обработкуперсональных данных в соответствии с политикой конфиденциальности
сульфат — цена анализа в Днипро в ИНВИТРО
Исследуемый материал Сыворотка крови
Метод определения Твёрдофазный хемилюминесцентный иммуноанализ
Половой гормон дегидроэпиандростерон сульфат является неактивной формой основного предшественника андрогена (ДГЭА), он содержится в крови и мужчин, и женщин. При половом созревании гормон играет немаловажную роль в формировании вторичных половых мужских признаков и способен преобразовываться в эстрадиол и тестостерон.
При половом созревании гормон играет немаловажную роль в формировании вторичных половых мужских признаков и способен преобразовываться в эстрадиол и тестостерон.
Дегидроэпиандростерон сульфат (ДГАЭС) продуцируется в надпочечниках, присоединяющийся к нему сульфат делает его «резервным», то есть биологически неактивным. Если же организм ощущает потребность в андрогенах, то сульфатная группа отщепляется, после чего активность гормона восстанавливается.
Анализ dhea s используют в следующих случаях:
- Для определения функции надпочечников.
- Для уточнения диагноза болезней, связанных с надпочечными железами и болезней, вызываемых нарушениями функций тестикул и яичников.
- Для помощи диагностики рака надпочечников, опухолей коры надпочечников, приобретенной или врожденной гиперплазии надпочечников.
- Для дополнительной диагностики синдрома поликистозных яичников.
- Для установления причин аменореи, бесплодия и гирсутизма.
- Для уточнения причин и диагностики маскулинизации у девочек и преждевременного полового созревания мальчиков.

У женщин избыточное количество андрогенов диагностируется как «гиперандрогения». Нарушенная выработка гормонов может стать причиной необратимых процессов, ведущих к образованию небольших кист и появлению плотной оболочки яичников. Это является одной из причин бесплодия.
Очень важна норма количества андрогенов при беременности. Для ее определения используется данный анализ. Если этот гормон в крови женщины повышен, необходимо срочно принимать соответствующие меры, так как это признак угрозы беременности. Своевременное лечение позволит будущей маме выносить и родить здоровенького ребенка.
Уровень ДГАЭС будет понижен, если отмечается задержание полового развития. Ряд исследований показали, что низкая концентрация гормона связана с риском ишемической болезни сердца.
Назначает анализ и делает расшифровку результатов определенный специалист. Исследование выполнит лаборатория ИНВИТРО, которая располагает лучшим оборудованием и предлагает приемлемые цены.
Перед сдачей анализа необходимо прекратить прием тиреоидных и стероидных гормонов, исключить и физические, и эмоциональные перенапряжения, не курить и не есть за два-три часа до исследования.
Дегидроэпиандростерон-сульфат (ДГЭА сульфат)
Дегидроэпиандростерон-сульфат (ДГЭА сульфат)
ДГЭА-С (дегидроэпиандростерон-сульфат) – основной стероид, секретируемый корой надпочечников (95%) и яичниками (5%). Выделяется с мочой и составляет основную фракцию 17-кетостероидов. В процессе его метаболизма в периферических тканях образуются тестостерон и дигидротестостерон. ДГЭА-С обладает относительно слабой андрогенной активностью, однако его биологическая активность усиливается благодаря относительно высоким концентрациям в сыворотке. Содержание ДГЭА-С в сыворотке является маркером синтеза андрогенов надпочечниками. Поскольку лишь незначительная часть гормона образуется половыми железами, определение концентрации ДГЭА-С может помочь в определении источника андрогенов. Если у женщины наблюдается повышенный уровень тестостерона, то с помощью определения концентрации ДГЭА-С можно установить, связано это с заболеванием надпочечников или яичников. В качестве маркера продукции андрогенов надпочечниками в настоящее время предпочитают определять ДГЭА-С в крови, а не 17-кетостероиды (17-КС) в моче.
В качестве маркера продукции андрогенов надпочечниками в настоящее время предпочитают определять ДГЭА-С в крови, а не 17-кетостероиды (17-КС) в моче.
Подготовка к исследованию
Исследование проводится утром, натощак. С последнего приема пищи должно пройти не менее 8 часов. Прекратить прием стероидных и тиреоидных гормонов за 48 часов до исследования (по согласованию с врачом).
Показания к исследованию
Для оценки функции надпочечников.
Для того чтобы отличить болезни, связанные с надпочечными железами, от заболеваний, вызванных нарушением функций яичников и тестикул.
Для помощи в диагностике опухолей коры надпочечников, рака надпочечников, врожденной либо приобретенной гиперплазии надпочечников и для отличия этих заболеваний от опухолей и рака яичников.
Для помощи в диагностике синдрома поликистозных яичников.
Для выяснения причин бесплодия, аменореи и гирсутизма.
Для диагностики и выяснения причин маскулинизации у девочек, а также преждевременного полового созревания у мальчиков.
Интерпретация
Референсные значения: мкг/дл.
|
Норма |
норма |
|
< 1недели |
108:607 |
|
1-4 недели |
31.6:431 |
|
1 месяц — 1 год |
3.4:124 |
|
1-5 лет |
0.47:19.4 |
|
5-10 лет |
2. |
|
10-15 лет |
33.9:280 |
|
15-20 лет |
65.1:368 |
|
20-25 лет |
148:407 |
|
25-35 лет |
98.8:340 |
|
35-45 лет |
60.9:337 |
|
45-55 лет |
35.4:256 |
|
55-65 лет |
18.9:205 |
|
65-75 лет |
9. |
|
>75 лет |
12:154 |
|
10-15 лет |
24.4:247 |
|
15-20 лет |
70.2:492 |
|
20-25 лет |
211:492 |
|
25-35 лет |
160:449 |
|
35-45 лет |
88.9:427 |
|
44-55 лет |
44.3:331 |
|
55-65 лет |
51. |
|
65-75 лет |
33.6:249 |
|
>75 лет |
16.2:123 |
 8:85.2
8:85.2 4:246
4:246 7:225
7:225Повышенный уровень ДГЭА-SO4 может свидетельствовать об опухоли коры надпочечников, раке либо гиперплазии надпочечников. Он не является диагностическим признаком какой-то определенной болезни, однако указывает на необходимость дальнейшего обследования для выявления причин гормонального дисбаланса.
Низкий уровень ДГЭА-SO4 бывает вызван дисфункцией надпочечников или гипопитуитаризмом, нарушениями, являющимися причиной понижения уровня питуитарных гормонов, которые, в свою очередь, регулируют производство и секрецию гормонов, производимых надпочечниками.
Назначается в комплексе с
Тестостерон
Фолликулостимулирующий гормон (ФСГ)
Лютеинизирующий гормон (ЛГ)
Пролактин
Глобулин, связывающий половые гормоны (ГСПГ)
Молекулярные основы распознавания лутропина рецептором маннозы / GalNAc-4-SO4
Реферат
Период полувыведения гликопротеинового гормона лутропина (ЛГ) из кровотока точно регулируется рецептором маннозы (Man) / GalNAc-4-SO 4 , экспрессируемым в эндотелиальных клетках печени. Быстрое выведение из кровотока способствует эпизодическому повышению и падению уровней ЛГ, что важно для максимальной стимуляции рецептора ЛГ, связанного с G-белком. Мы определили две молекулярные формы рецептора Man / GalNAc-4-SO 4 , которые различаются лигандной специфичностью, экспрессией в клетках и тканях и функцией.Форма, экспрессируемая эндотелиальными клетками печени, связывает лиганды, несущие GalNAc-4-SO 4 , и регулирует период полужизни гормона в кровообращении, тогда как форма, экспрессируемая макрофагами, связывает лиганды, несущие человека, и может играть роль в врожденном иммунитете. Мы демонстрируем, что GalNAc-4-SO 4 -специфическая форма в эндотелиальных клетках печени является димерной, тогда как Man-специфическая форма в макрофагах легких является мономерной, что объясняет различные лигандные специфичности рецептора, экспрессируемого в этих тканях.Два богатых цистеином домена, каждый из которых связывает один GalNAc-4-SO 4 , необходимы для образования стабильных комплексов с LH.
Быстрое выведение из кровотока способствует эпизодическому повышению и падению уровней ЛГ, что важно для максимальной стимуляции рецептора ЛГ, связанного с G-белком. Мы определили две молекулярные формы рецептора Man / GalNAc-4-SO 4 , которые различаются лигандной специфичностью, экспрессией в клетках и тканях и функцией.Форма, экспрессируемая эндотелиальными клетками печени, связывает лиганды, несущие GalNAc-4-SO 4 , и регулирует период полужизни гормона в кровообращении, тогда как форма, экспрессируемая макрофагами, связывает лиганды, несущие человека, и может играть роль в врожденном иммунитете. Мы демонстрируем, что GalNAc-4-SO 4 -специфическая форма в эндотелиальных клетках печени является димерной, тогда как Man-специфическая форма в макрофагах легких является мономерной, что объясняет различные лигандные специфичности рецептора, экспрессируемого в этих тканях.Два богатых цистеином домена, каждый из которых связывает один GalNAc-4-SO 4 , необходимы для образования стабильных комплексов с LH./65/65.jpg) Кинетика связывания ЛГ GalNAc-4-SO 4 -специфической формой рецептора в сочетании со скоростью его интернализации с поверхности клетки делает вероятным, что только два из четырех концевых GalNAc-4-SO 4 фрагменты, присутствующие в нативном ЛГ, задействуются до интернализации рецептора. В результате скорость клиренса гормона будет оставаться постоянной в широком диапазоне концентраций ЛГ и не будет чувствительна к изменениям количества концевых остатков GalNAc-4-SO 4 , пока два или более присутствуют на нескольких олигосахариды.
Кинетика связывания ЛГ GalNAc-4-SO 4 -специфической формой рецептора в сочетании со скоростью его интернализации с поверхности клетки делает вероятным, что только два из четырех концевых GalNAc-4-SO 4 фрагменты, присутствующие в нативном ЛГ, задействуются до интернализации рецептора. В результате скорость клиренса гормона будет оставаться постоянной в широком диапазоне концентраций ЛГ и не будет чувствительна к изменениям количества концевых остатков GalNAc-4-SO 4 , пока два или более присутствуют на нескольких олигосахариды.
Гликопротеиновые гормоны лутропин (ЛГ) и тиреотропин (ТТГ) несут N-связанные олигосахариды, оканчивающиеся последовательностью SO 4 -4-GalNAcβ1,4GlcNAcβ1,2Manα (1–4). Ранее мы идентифицировали рецептор, расположенный в эндотелиальных клетках печени, который распознает сульфатированные олигосахариды на ЛГ и опосредует быстрое удаление ЛГ из кровотока (5, 6). Экспрессия трансфераз, ответственных за синтез этих сульфатированных олигосахаридов, регулируется эстрогеном (7).Эти эволюционно консервативные уникальные сульфатированные структуры обнаружены на гликопротеиновых гормонах всех видов позвоночных (8). Регулируемое высвобождение ЛГ гонадотрофами в передней доле гипофиза в сочетании с его коротким периодом полужизни в кровотоке объясняют эпизодические подъемы и падения уровней гормонов, которые необходимы для оптимальной активации рецептора ЛГ, рецептора, сопряженного с G-белком (6, 9, 10).
Мы выделили рецептор из печени, который отвечает за клиренс ЛГ путем связывания терминального GalNAc-4-SO 4 .Этот рецептор имеет такую же пептидную карту, иммунореактивность и молекулярную массу, что и рецептор, специфичный для макрофагов маннозы (Man) (11). Поэтому мы назвали этот рецептор рецептором Man / GalNAc-4-SO 4 (12). Он состоит из богатого цистеином домена, повтора фибронектина типа II, восьми доменов узнавания углеводов C-типа (CRD), трансмембранного домена и цитозольного домена (13-15). Предыдущие исследования локализовали связывание человека, N- ацетилглюкозамина и фукозы с CRD 4-8 рецептора (16, 17).Напротив, мы локализовали связывание GalNAc-4-SO 4 с богатым Cys доменом (18). В отличие от рецептора макрофага Man из легких, который связывается с иммобилизованным человеком, но не с иммобилизованным LH, GalNAc-4-SO 4 -специфический рецептор, выделенный из печени, связывается с LH-сефарозой, но не с Man-сефарозой (11). Клетки яичника китайского хомячка (СНО), экспрессирующие рекомбинантный рецептор Man / GalNAc-4-SO 4 , связываются и интернализируют лиганды, несущие либо концевой Man, либо GalNAc-4-SO 4 , что позволяет предположить, что посттрансляционные различия наиболее вероятно объясняют Man- и GalNAc-4-SO 4 -специфические формы рецептора (12).
Различные лигандные специфичности, проявляемые рецептором Man / GalNAc-4-SO 4 , экспрессируемым макрофагами и эндотелиальными клетками печени, указывают на то, что рецептор используется для множества функций и что экспрессия этих специфичностей сильно регулируется. Поскольку существует только один богатый Cys домен, мы предположили, что рецептор Man / GalNAc-4-SO 4 должен быть димерным для связывания лигандов, несущих GalNAc-4-SO 4 . Образование димера может препятствовать доступу к сайтам внутри CRD 4-8, которые связывают человека.Напротив, мономерные формы рецептора Man / GalNAc-4-SO 4 д. Обеспечивать свободный доступ Man-несущих лигандов к CRD 4-8. Мы демонстрируем здесь, что GalNAc-4-SO 4 -специфическая форма рецептора действительно является димерной, тогда как Man-специфическая форма является мономерной. Кроме того, мы представляем доказательства того, что количество и расположение концевых остатков GalNAc-4-SO 4 на ЛГ имеет решающее значение для распознавания рецептором.
Материалы и методы
Трансфекций.
конструкций в pIg (19) котрансфицировали (6,25 мкг каждой кДНК) в клетки CHO / Tag 30A с использованием липофектамина (20). Через 48 часов после трансфекции клетки радиоактивно метили транс [ 35 S] -меткой (ICN) в течение 6 часов, и секретированные слитые белки выделяли из среды с помощью аффинной хроматографии на протеине A-сефарозе (18).
Анализ связывания с иммобилизованным человеком и ЛГ.
Очищенные химеры Fc инкубировали в течение ночи при 4 ° C с ман-сефарозой в 20 мМ трис-HCl (pH 7.4), 150 мМ NaCl (TBS), содержащий 2 мМ CaCl 2 . Те же химеры Fc инкубировали в LH-сефарозе в TBS, содержащем 5 мМ EDTA, для предотвращения связывания через Man-специфические сайты. Связанную химеру Fc элюировали с использованием 100 мМ глицина, pH 3,0 (18).
Описание рецептора Man / GalNAc-4-SO
4 , полученного из печени и легких.Man / GalNAc-4-SO 4 Рецептор из печени и легких крыс солюбилизировали в Triton X-100 и инкубировали с агглютинином зародышей пшеницы (WGA) -сефарозой в течение ночи при 4 ° C (11).После удаления несвязанных белков промыванием TBS + 0,1% Triton X-100 связанные гликопротеины элюировали TBS + 0,1% Triton X-100, содержащим 300 мМ GlcNAc. GlcNAc удаляли диализом против TBS + 0,1% Triton X-100. Аликвоты (100 мкл) материала, обогащенного WGA, инкубировали с 25 мкл LH в присутствии 5 мМ EDTA или 25 мкл Man-Sepharose в присутствии 5 мМ CaCl 2 для стимулирования связывания с Man-специфическими сайтами. Рецептор Man / GalNAc-4-SO 4 , который был связан либо с LH-сефарозой, либо с Man-сефарозой, элюировали 100 мМ глицином, pH 3.0, 0,1% Тритон Х-100.
Вестерн-блот анализов.
Элюированный Man / GalNAc-4-SO 4 Рецептор растворяли в буфере Лэммли при 25 ° C и разделяли невосстанавливающим SDS / PAGE (7,5% акриламида) (21) при 25 ° C. Белки электрофоретически переносили в Immobilon-P (Millipore) в 20 мМ Трис, 150 мМ глицине, 10% метаноле. Перенесенный рецептор Man / GalNAc-4-SO 4 был обнаружен путем инкубации с моноспецифическим кроличьим рецептором против Man / GalNAc-4-SO 4 , полученным при иммунизации кроликов кДНК для нативного Man / GalNAc-4-SO 4 Рецептор в инъецированном pCDM8 i.м. в физиологическом растворе и бустинг через 4 недели 20 мкг очищенной растворимой химеры Man / GalNAc-4-SO 4 -HAHis6 в неполном адъюванте Фрейнда. Химера Man / GalNAc-4-SO 4 -HAHis6 была получена путем замены трансмембранного и цитозольного доменов рецептора Man / GalNAc-4-SO 4 последовательностью из гемагглютинина гриппа, за которой следовали шесть остатков гистидина (HAH6 ). Рецептор anti-Man / GalNAc-4-SO 4 использовали в разведении 1: 10 000 и детектировали с помощью хемилюминесценции согласно производителю (NEN).
Анализ поверхностного плазмонного резонанса.
Анализ поверхностного плазмонного резонанса выполняли на приборе Amersham Pharmacia BIAcore 2000. Очищенный рекомбинантный белок A (Pierce) в концентрации 100 мкг / мл в 10 мМ натрий-ацетатном буфере, pH 4,5 присоединяли к сенсорному чипу BIAcore CM5 с использованием набора для связывания аминов, предоставленного производителем. Химеры Fc, связанные с иммобилизованным белком А, не диссоциировали в ходе анализов связывания. Проточная ячейка, лишенная связанного белка, была использована в качестве контрольной ячейки для корректировки неспецифического связывания и изменений значений единиц ответа, вызванных компонентами буфера.Анализы проводили при 7 ° C и скорости потока 5 мкл / мин в TBS, содержащем 0,005% поверхностно-активного вещества P20 и 5 мМ EDTA. Для измерения аффинности разные концентрации бычьего LH, LHα или LHβ (поставляемые Национальным институтом диабета, болезней пищеварения и почек Национальной гормональной и гипофизарной программой и AF Parlow) связывали в течение 10 минут (50 мкл), чтобы обеспечить достаточное количество время достичь равновесия. Связанные лиганды элюировали путем инъекции импульса 25 мкл 10 мМ Hepes (pH 7,4), 150 мМ NaCl, 3 мМ EDTA и 0.005% ПАВ Р20. Hepes ( N -2-гидроксиэтилпиперазин- N ‘-2-этансульфоновая кислота) имеет K i = 2,32 × 10 -4 M из-за сульфоната. Контрольные эксперименты проводили в связывающем буфере, содержащем 1 мМ GalNAc-4-SO 4 , который количественно ингибирует связывание богатым Cys доменом рецептора Man / GalNAc-4-SO 4 . Кривые насыщения анализировали с помощью нелинейной регрессии (программное обеспечение призмы, V2.0). Для конкурентных исследований возрастающие концентрации ингибиторов добавляли к фиксированной концентрации LHα (685 нМ).
Результаты
Химерные формы рецептора Man / GalNAc-4-SO
4 требуют двух богатых Cys домена для связывания иммобилизованного ЛГ.Мы использовали химерные формы рецептора Man / GalNAc-4-SO 4 , в которых трансмембранный и цитозольный домены рецептора были заменены Fc-областью человеческого IgG1, чтобы продемонстрировать, что богатый Cys домен отвечает за связывание терминал GalNAc-4-SO 4 (18). Химера Man / GalNAc-4-SO 4 -Fc (Mu1), которая содержит только CRD 1-8, связывает лиганды с концевым Man, но не лиганды с концевым GalNAc-4-SO 4 .Напротив, химера Fc Man / GalNAc-4-SO 4 -Fc (Mu11), состоящая из богатого Cys домена, связывает лиганды с концевым GalNAc-4-SO 4 , но не с концевым Man. Обе химеры Fc образуют ковалентные димеры, которые эффективно секретируются в среду при экспрессии в клетках СНО.
Коэкспрессия кДНК, кодирующих Mu1 и Mu11, в клетках CHO приводит к синтезу и секреции трех различных димерных видов. Большинство видов состоит из ( i ) ковалентно связанного гетеродимера, состоящего из одной цепи Mu1 и одной цепи Mu11, обозначенного Mu1 / 11, и ( ii ) гомодимера Mu11.Также были выражены следовые количества гомодимера Mu1. Гомодимерные и гетеродимерные химеры Fc легко идентифицируются при разделении невосстанавливающим SDS / PAGE (рис. 1). После выделения с помощью аффинной хроматографии на протеин А-сефарозе химеры, меченные 35 S-Cys / Met, инкубировали либо с ман-сефарозой, либо с LH-сефарозой. Количество каждой химеры в несвязанной и связанной фракциях определяли невосстанавливающим SDS / PAGE с последующей авторадиографией (рис. 1). Mu1 / 11 эффективно связывается с Man-Sepharose, но не с LH-Sepharose.Напротив, Mu11 связывается с LH-сефарозой, но не с Man-Sepharose. Таким образом, химеры Fc с одним богатым Cys доменом неспособны связываться с олигосахаридами, несущими GalNAc-4-SO 4 , на иммобилизованном ЛГ с достаточной стабильностью, чтобы сделать возможным их выделение. Напротив, химеры Fc с двумя богатыми Cys доменами способны связываться с иммобилизованным LH.
Рисунок 1Два богатых Cys домена необходимы для связывания с иммобилизованным LH. Man / GalNAc-4-SO 4 -Fc (Mu1) и Man / GalNAc-4-SO 4 -Fc (Mu11) котрансфицировали в клетки CHO / Tag30A и метили транс [ 35 S] -меткой. .Секретированные химеры Fc выделяли аффинной хроматографией на протеине А. Химеры Fc инкубировали с ман-сефарозой в TBS, содержащем 5 мМ Ca 2+ , или с LH-сефарозой в TBS, содержащем 5 мМ EDTA. Исходный материал для каждой матричной (входной), несвязанной (UB) и связанной (B) фракций исследовали невосстанавливающим SDS / PAGE. Положения Man / GalNAc-4-SO 4 -Fc (Mu1) 340000 Да, Man / GalNAc-4-SO 4 -Fc (Mu1 / 11) 220 000 Да и Man / GalNAc-4-SO Показано 4 -Fc (Mu11) 98000 Да.
В других исследованиях (данные не представлены) мы охарактеризовали конструкцию Man / GalNAc-4-SO 4 -HAHis6, в которой трансмембранный и цитозольный домены рецептора Man / GalNAc-4-SO 4 были заменены на антигенная последовательность гемагглютинина гриппа (22), за которой следуют шесть остатков гистидина. Химера Man / GalNAc-4-SO 4 -HAHis6 мигрирует как мономер при исследовании с помощью невосстанавливающего SDS / PAGE или гель-фильтрации. Man / GalNAc-4-SO 4 -HAHis6 не связывается с LH-сефарозой, но связывается с Man-сефарозой.Таким образом, мономерные формы рецептора Man / GalNAc-4-SO 4 , содержащие все его внеклеточные домены (Cys-богатые, фибронектиновые повторы типа II и CRD 1-8), неспособны связываться с иммобилизованным LH. Свойства этих химерных молекул демонстрируют, что для образования стабильных комплексов с олигосахаридами, несущими GalNAc-4-SO 4 , на LH требуются два богатых Cys домена.
GalNAc-4-SO
4 -специфические и специфичные для человека формы рецептора Man / GalNAc-4-SO 4 Обнаружен рецептор in vivo являются димерными и мономерными соответственно.Предыдущие исследования показали, что нативные и рекомбинантные формы рецептора макрофага Man существуют в виде мономеров (17). Однако после солюбилизации в Triton X-100 мы обнаружили мономерный и димерный рецептор Man / GalNAc-4-SO 4 в препаратах печени, исследованных с помощью невосстанавливающего SDS / PAGE и вестерн-блот-анализа. Далее мы исследовали свойства солюбилизированного рецептора Man / GalNAc-4-SO 4 , полученного из печени и легких крыс после обогащения с помощью лектиновой аффинной хроматографии с использованием агглютинин-сефарозы зародышей пшеницы.Частично очищенный рецептор инкубировали либо с LH-сефарозой, либо с ман-сефарозой, и относительные количества димерных и мономерных форм рецептора, присутствующих в связанных фракциях, определяли невосстанавливающим SDS / PAGE (фиг. 2). Девяносто семь процентов рецептора Man / GalNAc-4-SO 4 , полученного из легких, связывает Man-Sepharose (рис. 2). Напротив, 62% рецептора Man / GalNAc-4-SO 4 , полученного из печени, связывается с LH-сефарозой и 38% — с Man-сефарозой (рис. 2). Девяносто пять процентов рецептора Man / GalNAc-4-SO 4 , который выделяется из легких и связывается с Man-Sepharose, мигрирует в виде мономера (рис.2 В ). Напротив, 34% рецептора Man / GalNAc-4-SO 4 из печени, который связывается с LH-сефарозой, мигрирует как димер, тогда как 66% мигрирует как мономер (рис. 2 C ). Доля рецептора Man / GalNAc-4-SO 4 , которая мигрирует в виде димера, в 10 раз больше для рецептора, который связывается с LH-сефарозой (несущей концевой GalNAc-4-SO 4 ) по сравнению с рецептором что связывается с Человеком-Сефарозой. Поскольку ожидается, что нековалентные гомодимеры рецепторов диссоциируют во время SDS / PAGE, а электрофоретический перенос на Immobilon-P будет менее эффективным для димерных, чем для мономерных форм рецептора, значение 34% является минимальной оценкой количества димерного Man / GalNAc. -4-SO 4 рецептора , присутствующего во фракции, связанной с LH.
Рисунок 2Димерные и мономерные формы рецептора Man / GalNAc-4-SO 4 обнаружены in vivo и связываются с иммобилизованным LH и Man соответственно . Равные аликвоты частично очищенного рецептора Man / GalNAc-4-SO 4 из печени и легких крысы инкубировали с LH-сефарозой и Man-сефарозой. Связанный рецептор Man / GalNAc-4-SO 4 элюировали при pH 3,0, разделяли невосстанавливающим SDS / PAGE и электрофоретически переносили в Immobilon-P для вестерн-блот-анализа с использованием моноспецифического кроличьего антитела против Man / GalNAc-4-SO 4 рецепторные антитела.( A ) Рецептор Man / GalNAc-4-SO 4 из легких и печени, связанный с LH-сефарозой и Man-сефарозой, соответственно. Количество (условные единицы) мономера (серый) и димера (черный) определяли количественно с помощью денситометрии для легкого ( B ) и печени ( C ).
Свойства нативной GalNAc-4-SO 4 -специфичной и Man-специфической форм рецептора Man / GalNAc-4-SO 4 отражают результаты, полученные с растворимыми химерами Fc, что позволяет предположить, что Man / GalNAc-4-SO 4 / GalNAc-4-SO 4 Рецептор должен образовывать нековалентный димер для связывания LH-сефарозы с достаточной стабильностью для выделения.Кроме того, отсутствие значительных количеств димера в той части рецептора печени, которая связывается с Man-Sepharose, указывает на то, что сайты связывания Man, расположенные в CRD 4-8 (16), изолированы в GalNAc-4-SO 4 — специфическая форма рецептора.
Количество и расположение терминала GalNAc-4-SO
4 частей имеют решающее значение для распознавания LH.Чтобы понять значение количества и расположения концевых остатков GalNAc-4-SO 4 , мы дополнительно охарактеризовали N-связанные олигосахариды на бычьем ЛГ, гетеродимер, состоящий из α-субъединицы и β-субъединицы (23), и его отдельные субъединицы.Олигосахариды на димере LHβ, LHα и LH высвобождались путем расщепления с помощью PNGase F и дериватизировались 9-аминопирен-1,4,6-трисульфоновой кислотой (Beckman) для анализа с помощью капиллярного электрофореза (24). Одиночный N-связанный олигосахарид на LHβ состоит исключительно (99%) из сложных двухантенарных структур с двумя концевыми остатками GalNAc-4-SO 4 (S-2), тогда как 90% олигосахаридов на двух сайтах N-связанного гликозилирования на LHα представляют собой сложные и гибридные структуры, каждая из которых содержит один концевой фрагмент GalNAc-4-SO 4 (S-1) (E.I. Park и J.U.B., неопубликованное наблюдение) (см. Схему на рис. 4). Распределение олигосахаридов на димере LH равно сумме обнаруженных на разделенных субъединицах α и β.
Мы использовали поверхностный плазмонный резонанс для оценки связывания химерой Mu11 Fc, которая имеет два богатых Cys домена. Белок А, иммобилизованный на чипах биосенсора, использовали для захвата Mu11. Стабильный комплекс способен взаимодействовать с концевыми остатками GalNAc-4-SO 4 на димере LHβ, LHα и LH.Кривые связывания были построены для LHβ (один S-2), LHα (два S-1) и димера LH (один S-2 и два S-1). Количество связанного в равновесии лиганда использовали для построения кривых насыщения, показанных на фиг. 3. Константы диссоциации и значения B max определяли с помощью нелинейного регрессионного анализа, как показано в таблице 1. LHβ связывается с Mu11 в концентрации -зависимым и насыщаемым способом с K d 2,06 × 10 −6 M (рис. 3 A и таблица 1).Кроме того, 1,12 моль LHβ связывается на моль димерного Mu11, что указывает на то, что каждый богатый Cys домен может взаимодействовать с одним остатком GalNAc-4-SO 4 . Это также демонстрирует, что два концевых фрагмента GalNAc-4-SO 4 на одном олигосахариде S-2 могут соединять участки связывания GalNAc-4-SO 4 на двух богатых Cys доменах, присутствующих в Fc-химере Mu11 . Однако LHα с двумя олигосахаридами S-1 проявляет в 10 раз лучшее сродство к Man / GalNAc-4-SO 4 -Fc (Mu11), чем LHβ с K d из 2.32 × 10 −7 M (рис.3 B , таблица 1 и схема на рис.4). Мольное соотношение, полученное с LHα, указывает на то, что один GalNAc-4-SO 4 из каждого из моносульфатированных олигосахаридов S-1 связан с каждым Cys-богатым доменом в Fc-химере. 10-кратное увеличение аффинности с LHα по сравнению с LHβ предполагает, что доступ концевых фрагментов GalNAc-4-SO 4 к сайту связывания на каждом Cys-богатом домене значительно лучше, когда GalNAc-4-SO 4 фрагментов присутствуют на отдельных олигосахаридных цепях.Сродство к LHα и LHβ отражает различия в пространственных отношениях концевых остатков GalNAc-4-SO 4 , поскольку они имеют одинаковые B max при насыщении, т. Е. Один связанный GalNAc-4-SO 4 на Cys-богатый домен. Расстояние между фрагментами GalNAc-4-SO 4 в двухантенарной структуре S-2 на LHβ может быть менее оптимальным или напряженным. Пространственные отношения концевых фрагментов GalNAc-4-SO 4 в структурах S-1 и S-2 диктуются расположением этих олигосахаридов на лежащих в основе пептидах субъединиц LHα и LHβ.
Рисунок 3Анализ связывания LHβ ( A ), LHα ( B ) и димера LH ( C ) химерой Man / GalNAc-4-SO 4 — Fc (Mu11) с использованием поверхностного плазмонного резонанса . Химера Man / GalNAc-4-SO 4 -Fc (Mu11) была иммобилизована на протеине-A, связанном с чипом CM5 Biacore. Увеличивающимся концентрациям димеров LHβ, LHα и LH позволяли связываться с химерой. Нелинейный регрессионный анализ полученных кривых насыщения проводился с использованием программного обеспечения призмы.Графики Скэтчарда также были построены и показаны на вставках. N — мольное отношение связанного лиганда на моль химеры Mu11 Fc, а F — концентрация лиганда. Мольные отношения определяли путем деления полученных значений единиц отклика на относительные молекулярные массы 98 128 для Man / GalNAc-4-SO 4 -Fc (Mu11), 14 880 для LHβ, 14 607 для LHα и 29 500 для димера LH.
Таблица 1Связывающие свойства лигандов с различным числом концевых GalNAc-4-SO 4 -частей для Man / GalNAc-4-SO 4 -Fc (Mu11)
Рисунок 4Модель для Man-специфической и GalNAc-4-SO 4 — специфической формы рецептора Man / GalNAc-4-SO 4 .Макрофаги экспрессируют преимущественно мономерную форму рецептора Man / GalNAc-4-SO 4 ( Left ), показанного встроенным в липидный бислой. Man-специфические CRD 4-8 (темные квадраты) доступны для Man-несущих лигандов, тогда как мономерный Cys-богатый домен (кружки) не способен образовывать стабильные комплексы с GalNAc-4-SO 4 -несущими лигандами. Эндотелиальные клетки печени экспрессируют преимущественно димерную форму рецептора Man / GalNAc-4-SO 4 ( Right ).Сопоставление двух Cys-богатых доменов позволяет формировать стабильные комплексы с лигандами, такими как LH, несущими два или более олигосахаридов с концевыми частями GalNAc-4-SO 4 ( верхний правый ). Показано, что рецептор взаимодействует с двумя структурами S-1 на LHα. Другие комбинации структур S-1 и S-2 могут дать такой же результат. Сайты связывания Man, обнаруженные в CRD 4-8, однако, изолированы от Man-несущих лигандов в димерной форме рецептора.
В отличие от LHα и LHβ, которые оба имеют один аффинный сайт для связывания, димер LH формирует кривую линейную диаграмму Скэтчарда (рис.3 C ), состоящий из двух или более компонентов. Регрессионный анализ дает один компонент с K d 2,67 × 10 -7 M, который по существу идентичен таковому, полученному с LHα, что указывает на то, что два из четырех концевых остатков GalNAc-4-SO 4 на Димер LH задействован. Дополнительный компонент с K d из 8,32 × 10 −9 M предполагает участие 3-4 частей концевого GalNAc-4-SO 4 в димере LH при связывании с химерой Fc Иммобилизованный Mu11 на чипе Biacore.
Ингибирование связывания LHα с Mu11 за счет увеличения концентраций GalNAc-4-SO 4 и SO 4 -4-GalNAcβ1,4GlcNAcβ1,2Manα-MCO контролировали с помощью поверхностного плазмонного резонанса (Таблица 1). GalNAc-4-SO 4 и SO 4 -4-GalNAcβ1,4GlcNAcβ1,2Manα-MCO имеют K i значения 16,8 × 10 -5 M и 1,20 × 10 -5 M, соответственно. Таким образом, богатый Cys домен распознает почти исключительно концевой сульфатированный моносахарид.Основная структура олигосахарида не вносит значительного вклада в связывание концевого GalNAc-4-SO 4 .
Обсуждение
Быстрый выведение нативного (сульфатированного), но не рекомбинантного (сиалилированного) бычьего ЛГ из кровотока привело нас к идентификации рецептора, специфичного для терминального GalNAc-4-SO 4 (6) (см. Рис. 4). Мы выделили рецептор, ответственный за выведение ЛГ из кровотока эндотелиальными клетками печени (5).Каждая эндотелиальная клетка способна связывать 5,7 × 10 5 молекул ЛГ с K d 1,63 × 10 -7 M (5). Кроме того, рецептор различает SO 4 -4-GalNAcβ1,4GlcNAcβ1,2Manα-BSA и SO 4 -3-GalNAcβ1,4GlcNAcβ1,2Manα-BSA, что указывает на то, что положение концевого сульфата имеет решающее значение для связывания и клиренса. из тиража (5).
GalNAc-4-SO 4 -специфический рецептор был выделен из печени крысы и, как было установлено, имеет ту же аминокислотную последовательность, что и Man-специфический рецептор, выделенный из альвеолярных макрофагов (11).Две формы рецептора различаются по своим свойствам связывания лиганда. Специфическая для GalNAc-4-SO 4 форма не связывается с иммобилизованным человеком, тогда как специфическая для человека форма не связывает иммобилизованные гликопротеины, несущие олигосахариды, оканчивающиеся на GalNAc-4-SO 4 . Однако клетки СНО могут экспрессировать рекомбинантные рецепторы, которые связывают и интернализируют гликопротеины, несущие либо концевой Man, либо концевой GalNAc-4-SO 4 (12). Это предполагает, что посттрансляционные события регулируют специфичность рецептора Man / GalNAc-4-SO 4 .Различная специфичность двух форм рецептора Man / GalNAc-4-SO 4 дополнительно указывает на то, что этот рецептор выполняет несколько функций. Эти функции отражают как лигандную специфичность рецептора, так и его сайт-специфическую экспрессию в разных клетках. Рецептор в эндотелиальных клетках печени, который удаляет из крови лиганды, несущие GalNAc-4-SO 4 , должен иметь одну или несколько структурных особенностей, которые отличают его от того же рецептора, экспрессируемого в макрофагах, который связывает лиганды с конечным человеком.
Основываясь на свойствах рецептора in vivo и свойств Fc-химер, мы предполагаем, что рецептор Man / GalNAc-4-SO 4 существует как в виде мономера, так и в виде гомодимера. Как показано на рис. 4, наши данные показывают, что мономерная форма рецептора Man / GalNAc-4-SO 4 является Man-специфической, а димерная форма является GalNAc-4-SO 4 -специфичной. Как Man-специфическая, так и GalNAc-4-SO 4 -специфическая формы рецептора присутствуют в эндотелиальных клетках печени, что позволяет предположить, что они находятся в равновесии друг с другом.Напротив, мы обнаруживаем только Man-специфическую форму в альвеолярных макрофагах, что согласуется с возможностью того, что рецептор Man / GalNAc-4-SO 4 в макрофагах не может образовывать гомодимер in vivo . Различные паттерны гликозилирования или других посттрансляционных модификаций в макрофагах и эндотелиальных клетках печени могут определять, способен ли рецептор образовывать димер in vivo . Различия в подвижности рецептора, выделенного из печени и легких (рис.2) согласуются с этой возможностью. В настоящее время мы исследуем молекулярную основу, определяющую, какая форма рецептора Man / GalNAc-4-SO 4 экспрессируется конкретной клеткой.
Наши предыдущие исследования продемонстрировали, что богатый Cys домен необходим и достаточен для связывания GalNAc-4-SO 4 , тогда как CRD 4-8 отвечают за связывание лигандов, заканчивающихся Man, GlcNAc или фукозой (18). Модель на рис.4 обеспечивает структурную основу для различий между GalNAc-4-SO 4 — и специфическими для человека формами рецептора, которые могут объяснить тот факт, что области, связывающие GalNAc-4-SO 4 и Человек физически различны.Текущие исследования показывают, что для образования стабильных комплексов с иммобилизованными лигандами, несущими несколько концевых частей GalNAc-4-SO 4 , требуются два богатых Cys домена. Кроме того, ≥62% рецептора Man / GalNAc-4-SO 4 в эндотелиальных клетках печени присутствует в форме гомодимера, поскольку мономерная форма не связывается с ЛГ.
Мы показали, что 93% рецептора Man / GalNAc-4-SO 4 в легких является мономерным на основании как его неспособности связываться с LH-сефарозой, так и отсутствия димерных частиц во время анализа с помощью SDS / PAGE.Доминирование мономерного рецептора в легких поддерживает наше предположение о том, что рецептор, экспрессируемый макрофагами, не может образовывать димерные виды. Более того, отсутствие димера в рецепторе, связанном с Man-Sepharose из печени, указывает на то, что CRD 4-8 действительно секвестированы в димерной форме и препятствуют доступу к иммобилизованным остаткам Man. Мономерные виды в печени могут отражать либо присутствие рецептора, который не может образовывать гомодимеры, либо мономеры, находящиеся в равновесии с димерами.
Рецептор Man / GalNAc-4-SO 4 примечателен тем, что его две специфичности в отношении лигандов, несущих GalNAc-4-SO 4 , и лигандов, несущих человек, являются взаимоисключающими. Доминирование мономерного рецептора в макрофаге, где доступны CRD 4-8, может отражать его предполагаемую роль в качестве фагоцитарного рецептора в врожденном иммунитете (25). Димерный GalNAc-4-SO 4 -специфический рецептор в эндотелиальных клетках необходим для контроля уровней ЛГ и ТТГ в кровотоке (5, 6, 26).Присутствие этих альтернативных форм в различных тканях указывает на то, что экспрессия регулируется клеточно-специфическим образом.
Гликопротеиновые гормоны ЛГ и ТТГ в настоящее время являются единственными хорошо зарекомендовавшими себя нативными лигандами для GalNAc-4-SO 4 -специфической формы рецептора Man / GalNAc-4-SO 4 . Оба гормона несут несколько олигосахаридов, заканчивающихся одним или несколькими фрагментами GalNAc-4-SO 4 . Их пространственные отношения имеют решающее значение для связывания, потому что присутствие двух олигосахаридов S-1 позволяет LHα связываться с K d из 2.32 × 10 -7 M, тогда как единственный олигосахарид S-2 на LHβ связывается с аффинностью в 10 раз ниже. Стехиометрия связывания указывает на то, что каждый богатый Cys доменом взаимодействует с одним GalNAc-4-SO 4 . Это согласуется с недавним кристаллографическим исследованием, в котором был идентифицирован единственный сайт связывания (27), и с исследованиями равновесного диализа (неопубликованная работа). Очевидно, что взаимосвязь концевых остатков GalNAc-4-SO 4 в пространстве оказывает большое влияние на их способность связываться одновременно с двумя богатыми Cys доменами.Наши данные показывают, что только определенные формы нативного рецептора Man / GalNAc-4-SO 4 , такие как тот, который присутствует в эндотелиальных клетках печени, образуют стабильные димерные структуры.
LH связывается изолированными эндотелиальными клетками печени с кажущимся K d 1,63 × 10 −7 M (5), значение почти идентично K d 2,67 × 10 −7. M получено для связывания LHα через две концевые группы GalNAc-4-SO 4 на отдельных олигосахаридах с химерой Fc, иммобилизованной на чипе Biacore.Это предполагает, что ЛГ связывается с димерной, GalNAc-4-SO 4 -специфической формой рецептора Man / GalNAc-4-SO 4 на поверхности эндотелиальных клеток, используя только два из его 3-4-концевых GalNAc- 4-СО 4 -множеств. Даже несмотря на то, что димер LH может связываться с рекомбинантной химерой Mu11, иммобилизованной на чипе Biacore с дополнительным компонентом K d 8,3 × 10 −9 M при взаимодействии с 3–4-концевым GalNAc-4-SO 4 фрагментов, этот последний компонент с высоким сродством не наблюдается, когда LH связывается с рецептором на поверхности эндотелиальных клеток печени in vivo .
На основании исследований связывания, проведенных с рекомбинантным рецептором Simpson et al. (28) предположили, что гликопротеиновые гормоны также могут связываться с рецептором Man / GalNAc-4-SO 4 через остатки Man отдельно или в комбинации с GalNAc-4-SO 4 . Бычий ЛГ действительно содержит концевые остатки Man, что обусловлено наличием гибридных структур (1, 2). Simpson et al. (28) сообщил, что ЛГ связывается с Man-специфическими сайтами рецептора Man / GalNAc-4-SO 4 со сродством в диапазоне 2.8 × 10 −6 M (28). В свете K d 2,32 × 10 -7 M, полученного при взаимодействии двух фрагментов GalNAc-4-SO 4 , кажется маловероятным, что взаимодействие с концевыми остатками Man способствует клиренсу LH в vivo . Секвестрация Man-связывающих доменов в димерной, GalNAc-4-SO 4 -специфической форме рецептора делает взаимодействие с концевыми остатками Man на LH еще менее вероятным. Способность нативного рецептора GalNAc-4-SO 4 из печени связывать ЛГ не изменяется присутствием человека, и мы не смогли продемонстрировать одновременное взаимодействие с человеком и GalNAc-4-SO 4 для любого человека. рецептор из легких или рецептор GalNAc-4-SO 4 из печени (D.Fiete and J.U.B., неопубликованное наблюдение). ЛГ и ТТГ от других видов животных не несут терминальный Man или GlcNAc в той же степени, что и бычий гормон, хотя они несут терминальный GalNAc-4-SO 4 (1, 2). Напротив, присутствие концевого β1,4-связанного GalNAc-4-SO 4 на N-связанных олигосахаридах гликопротеиновых гормонов было законсервировано у всех видов позвоночных, что указывает на критическую роль этих структур in vivo (8 ). Несмотря на то, что одновременное связывание с концевым Man и GalNAc-4-SO 4 на бычьем ЛГ при некоторых обстоятельствах возможно, эта форма связывания также вряд ли будет способствовать клиренсу ЛГ и ТТГ печеночным Man / GalNAc-4- SO 4 рецептор in vivo .
Наши результаты показывают, что форма рецептора Man / GalNAc-4-SO 4 , экспрессируемого эндотелиальными клетками по сравнению с макрофагами, строго регулируется. Кроме того, мы предоставили молекулярную основу того, как количество концевых остатков GalNAc-4-SO 4 на LH и TSH контролирует их период полураспада в кровотоке. Присутствия двух или более концевых остатков GalNAc-4-SO 4 на отдельных олигосахаридах достаточно для обеспечения продуктивного связывания рецептором Man / GalNAc-4-SO 4 на эндотелиальных клетках печени.Поскольку рецептор является димерным на эндотелиальных клетках и постоянно интернализуется, дополнительные остатки концевых GalNAc-4-SO 4 , присутствующие на нативном LH, вряд ли будут способствовать связыванию in vivo . Скорость ассоциации и концентрация ЛГ в крови будут определять, какая часть рецептора занята во время интернализации. Большое количество рецептора, экспрессируемого эндотелиальными клетками печени, гарантирует, что скорость выведения из кровотока за счет интернализации будет оставаться постоянной в широком диапазоне концентраций ЛГ, обеспечивая максимальную активацию рецептора ЛГ в яичниках и семенниках.
Благодарности
Мы благодарим Нэнси Баензигер, Джереми Кеуша и Элисон Вудворт за критические комментарии. Работа поддержана грантами R37-CA21923 и R01-DK41738 Национальных институтов здравоохранения.
Сноски
↵ * Кому следует обращаться с запросами на перепечатку. Электронная почта: Baenziger {at} Pathology.wustl.edu.
Этот документ был отправлен напрямую (Трек II) в офис PNAS.
Статья перед печатью опубликована на сайте: Proc. Natl. Акад. Sci. USA , 10.1073 / pnas.170184597.
Статья и дата публикации находятся на сайте www.pnas.org/cgi/doi/10.1073/pnas.170184597
Сокращения
- CHO,
- яичник китайского хомячка;
- Man,
- манноза;
- LH,
- лутропин;
- ФСГ,
- фоллитропин;
- ТТГ,
- тиреотропин;
- CRD,
- домен распознавания углеводов C-типа
- Copyright © Национальная академия наук
Арбускулярный микоризный симбиоз вызывает протеомные реакции, противоположные P-голоданию в подвое виноградной лозы SO4 при колонизации корней двумя видами Glomus
Abel S, Ticconi CA, Delatorre CA (2002) Фосфатное зондирование у высших растений.Physiol Plant 115: 1–8
PubMed CAS Google Scholar
Агуин О., Мансилла Дж. П., Виларино А., Сайнс М. Дж. (2004) Влияние микоризной инокуляции на морфологию корней и производство в питомниках трех подвоев виноградной лозы. Амер Дж. Энол Витикульт 55: 108–111
Google Scholar
Aloui A, Recorbet G, Gollotte A, Robert F, Valot B, Gianinazzi-Pearson V, Aschi-Smiti S, Dumas-Gaudot E (2009) О механизмах снятия напряжения кадмия в Medicago truncatula by arbuscular микоризный симбиоз: корневое протеомное исследование.Протеомика 9: 420–433
PubMed CAS Google Scholar
Андреадели А., Флеметакис Э., Аксарли И., Димоу М., Удварди М.К., Катинакис П., Лабру Н.Э. (2009) Клонирование и характеристика формиатдегидрогеназы Lotus japonicus : возможная корреляция с гипоксией. Biochim Biophys Acta 1794: 976–984
PubMed CAS Google Scholar
Azcón-Aguilar C, Barea JM (1996) Арбускулярная микориза и биологический контроль переносимых через почву патогенов растений — обзор задействованных механизмов.Микориза 6: 457–464
Google Scholar
Basha SM, Mazhar H, Vasanthaiah HKN (2010) Протеомический подход к идентификации уникальных белков ксилемного сока у устойчивых к болезни видов Vitis Пирса. Appl Biochem Biotech 160: 932–944
CAS Google Scholar
Balestrini R, Lanfranco L (2006) Экспрессия генов грибов и растений в симбиозе арбускулярной микоризы.Микориза 16: 509–524
PubMed CAS Google Scholar
Bavaresco L, Fogher C (1992) Эффект заражения корней Pseudomonas fluorescens и Glomus mosseae на повышение Fe-эффективности непривитых подвоев виноградной лозы. Vitis 31: 163–168
CAS Google Scholar
Bavaresco L, Fogher C (1996) Лайм-индуцированный хлороз виноградной лозы, пораженный подвой и заражением корней арбускулярной микоризой и Pseudomonas fluorescens .Vitis 35: 119–123
CAS Google Scholar
Берта Г., Фускони А. (1997) Влияние арбускулярных микоризных и эрикоидных грибов на структуру и активность ядер клетки-хозяина. В: Bryant J, Chiatante D (eds) Растения и пролиферация клеток. Wiley, Chichester, pp 135–151
Google Scholar
Bestel-Corre G, Dumas-Gaudot E, Poinsot V, Dieu M, Dierick JF, van Tuinen D, Remacle J, Gianinazzi-Pearson V, Gianinazzi S (2002) Анализ протеома и идентификация белков, связанных с симбиозом, из Medicago truncatula Gaertn методом двумерного электрофореза и масс-спектрометрии.Электрофорез 23: 122–137
PubMed CAS Google Scholar
Bestel-Corre G, Gianinazzi S, Dumas-Gaudot E (2004) Воздействие осадков сточных вод на симбиотические протеомы Medicago truncatula . Фитохимия 65: 1651–1659
PubMed CAS Google Scholar
Branscheid A, Sieh D, Pant BD, May P, Devers EA, Elkrog A, Schauser L, Scheible WR, Krajinski F (2010) Характер экспрессии предполагает роль MiR399 в регуляции клеточного ответа на локальный Pi увеличивается при арбускулярном микоризном симбиозе.Mol Plant Microbe Interact 23: 915–926
PubMed CAS Google Scholar
Brechenmacher L, Weidmann S, van Tuinen D, Chatagnier O, Gianinazzi S, Franken P, Gianinazzi-Pearson V (2004) Профилирование экспрессии активированных генов растений и грибов на ранних и поздних стадиях Medicago truncatula – Glomus mosseae взаимодействий. Микориза 14: 253–262
PubMed CAS Google Scholar
Bucher M (2007) Функциональная биология поглощения фосфатов растениями на границе корня и микоризы.New Phytol 173: 11–26
PubMed CAS Google Scholar
Camprubí A, Estaún V, Nogales A, García-Figueres F, Pitet M, Calvet C (2008) Реакция подвоя виноградной лозы Richter 110 на инокуляцию местными и отобранными арбускулярными микоризными грибами и показатели роста на пересадке виноградника. Микориза 18: 211–216. DOI: 10.1007 / s00572-008-0168-3
PubMed Google Scholar
Carvalho LC, Esquivel MG, Martins I, Ricardo CP, Amancio S (2005) Мониторинг стабильности Rubisco в микроклональном размножении виноградной лозы ( Vitis vinifera L.) методом двумерного электрофореза. J Plant Physiol 162: 365–374
PubMed CAS Google Scholar
Castro AJ, Carapito C, Zorn N, Magne C, Leize E, Van Dorsselaer A, Clément C (2005) Протеомный анализ тканей виноградной лозы ( Vitis vinifera L.), подвергнутых гербицидному стрессу. J Exp Bot 56: 2783–2795
PubMed CAS Google Scholar
Chalker-Scott L (1999) Экологическое значение антоцианов в ответах растений на стресс.Photochem Photobiol 70: 1–9
CAS Google Scholar
Cheng XM, Baumgartner K (2004) Исследование сообществ арбускулярных микоризных грибов на виноградниках Северной Калифорнии и потенциал микоризной колонизации питомников виноградных лоз. Hort Sci 39: 1702–1706
Google Scholar
Cheng XM, Baumgartner K (2006) Влияние микоризных корней и экстрарадикальных гиф на поглощение 15N из подстилки покровных культур виноградников и микробного сообщества почвы.Soil Biol Biochem 38: 2665–2675
CAS Google Scholar
Chung HJ, Ferl RJ (1999) Экспрессия алкогольдегидрогеназы Arabidopsis как в побегах, так и в корнях обусловлена средой роста корней. Физиология растений 121: 429–436
PubMed CAS Google Scholar
Дегучи Й, Банба М, Симода Й, Чечитка С.А., Сузури Р., Окусако Й, Ясухиро Ооки, Коичи Тоёкура К., Судзуки А., Утиуми Т., Хигачи И., Абэ М., Коучи Х, Изуи К., Синго Хата С. (2007) Транскриптомное профилирование корней Lotus japonicus во время развития арбускулярной микоризы и сравнение с таковым при клубеньке.ДНК Res 14: 117–133
PubMed CAS Google Scholar
Dumas-Gaudot E, Gollotte A, Cordier C, Gianinazzi S, Gianinazzi-Pearson V (2000) Модуляция систем защиты хозяина. В: Капульник Ю., Дудс Д.Д. (ред.) Арбускулярная микориза: физиология и функция. Kluwer Academic, Dordrecht, стр. 173–200
Google Scholar
Дюма-Годо Э, Вало Б, Бестель-Корре Г, Рекорбет Г, СТ-Арно М, Фонтен Б, Дьё М., Раес М., Сараванан Р.С., Джанинацци С. (2004) Протеомика как способ выявления дополнительных корешковые белки грибов из Glomus intraradices — RiT-ДНК микоризы корня моркови.FEMS Microbiol Ecol 48: 401–411
PubMed CAS Google Scholar
Duff SMG, Sarath G, Plaxton WC (2006) Роль кислых фосфатаз в метаболизме фосфора растений. Physiol Plant 90: 791–800
Google Scholar
Fiorilli V, Catoni M, Miozzi L, Novero M, Accotto GP, Lanfranco L (2009) Глобальные профили экспрессии генов и генов клеточного типа в растениях томатов, колонизированных арбускулярным микоризным грибом.Новый Фитол. 184: 975–987. DOI: 10.1111 / j.1469-8137.2009.03031.x
PubMed CAS Google Scholar
Franco-Zorrilla JM, González E, Bustos R, Linhares F, Leyva A, Paz-Ares J (2004) Транскрипционный контроль реакции растений на ограничение фосфата. J Exp Bot 55: 285–293
PubMed CAS Google Scholar
Гарсия-Гарридо Дж. М., Окампо Дж. А. (2002) Регулирование защитной реакции растений при арбускулярном микоризном симбиозе.J Exp Bot 53: 1377–1386
PubMed CAS Google Scholar
Genre A, Bonfante P (1998) Конфигурация актина и тубулина в арбускул-содержащих клетках из корней микоризного табака. Новый Фитол 140: 745–752
CAS Google Scholar
Giovannini E (1999) Produção de uvas para vinho, suco e mesa. Renascença, Порту-Алегри, стр. 364
Google Scholar
Görg A, Postel W, Weser J, Günther S, Strahler JR, Hanash SM, Somerlot L (1987) Устранение точечных штрихов на окрашенных серебром двумерных гелях путем добавления йодацетамида в буфер для уравновешивания.Электрофорез 8: 122–124
Google Scholar
Grunwald U, Nyamsuren O, Tamasloukht MB, Lapopin L, Becker A, Mann P, Gianinazzi-Pearson V, Krajinski F, Franken P (2004) Идентификация регулируемых микоризой генов с профилем экспрессии, связанным с развитием арбускул. Завод Мол Биол 55: 553–566
PubMed CAS Google Scholar
Guether M, Balestrini R, Hannah M, He J, Udvardi MK, Bonfante P (2009) Общегеномное перепрограммирование регуляторных сетей, транспорта, клеточной стенки и биогенеза мембран во время арбускулярного микоризного симбиоза в Lotus japonicus.Новый Фитол 182: 200–212
PubMed CAS Google Scholar
Güimil S, Chang HS, Zhu T, Sesma A, Osbourn A, Roux C, Ioannidis V, Oakeley EJ, Docquier M, Descombes P, Briggs SP, Paszkowski U (2005) Сравнительная транскриптомика риса показывает древний образец реакции на микробную колонизацию. Proc Natl Acad Sci USA 102: 8066–8070
PubMed Google Scholar
Harrier LA, Watson CA (2004) Потенциальная роль арбускулярных микоризных (AM) грибов в биозащите растений от почвенных патогенов в органических и / или других устойчивых сельскохозяйственных системах.Pest Manag Sci 60: 149–157
PubMed CAS Google Scholar
Харрисон MJ (1999) Молекулярные и клеточные аспекты арбускулярного микоризного симбиоза. Annu Rev Plant Physiol Plant Mol Bio 50: 361–389
CAS Google Scholar
Hewitt EJ (1966) Методы культивирования в песке и воде, используемые в исследованиях питания растений. Сельскохозяйственное бюро Содружества, Лондон
Google Scholar
Hohnjec N, Vieweg MF, Puhler A, Becker A, Kuster H (2005) Перекрытие транскрипционных профилей корней Medicago truncatula , инокулированных двумя разными грибами Glomus , дает представление о генетической программе, активируемой во время арбускулярной микоризы .Физиология растений 137: 1283–1301
PubMed CAS Google Scholar
Hourton-Cabassa C, Ambard-Bretteville F, Moreau F, Davy de Virville J, Remy R, des Francs-Small CC (1998) Стресс-индукция митохондриальной формиатдегидрогеназы в листьях картофеля. Физиология растений 116: 627–635
PubMed CAS Google Scholar
Ignoul S, Eggermont J (2005) Домены CBS: структура, функция и патология белков человека.Am J Physiol Cell Physiol 289: C1369 – C1378
PubMed CAS Google Scholar
Javot H, Pumplin N, Harrison MJ (2007) Фосфат в арбускулярном микоризном симбиозе: транспортные свойства и регуляторные роли. Среда растительной клетки 30: 310–322
PubMed CAS Google Scholar
Jaillon O, Aury JM, Noel B, Policriti A, Clepet C et al (2007) Последовательность генома виноградной лозы предполагает наследственную гексаплоидизацию в основных типах покрытосеменных.Nature 449: 463–467
PubMed CAS Google Scholar
Jiang J, Clouse SD (2001) Экспрессия гена растения со сходной последовательностью с белком, взаимодействующим с рецептором TGF-бета животного, регулируется брассиностероидами и необходима для нормального развития растений. Завод J 26: 35–45
PubMed CAS Google Scholar
Джонсон С.М., Ульрих А. (1959) Аналитические методы для использования в анализе растений.Бюллетень 766, Калифорнийская сельскохозяйственная экспериментальная станция, стр. 25–78
Карагианнидис Н., Николау Н., Маттеу А. (1995) Влияние трех видов VA-микоризы на рост и поглощение питательных веществ тремя подвоями виноградной лозы и одним столовым сортом винограда. Vitis 34: 85–89
Google Scholar
Karagiannidis N, Nikolaou N, Ipsilantis I, Zioziou E (2007) Влияние различных азотных удобрений на активность Glomus mosseae , а также на питание виноградной лозы и состав ягод.Микориза 18: 43–50. DOI: 10.1007 / s00572-007-0153-2
PubMed CAS Google Scholar
Krüger A, Peskan-Berghöfer T, Frettinger P, Herrmann S, Buscot F, Oelmüller R (2004) Идентификация генов растений, связанных с премикоризой, в ассоциации между Quercus robur и Piloderma croceum . Новый Фитол 163: 149–157
Google Scholar
Li K, Xu C, Zhang K, Yang A, Zhang J (2007) Протеомный анализ роста корней и метаболических изменений при дефиците фосфора у кукурузы ( Zea mays L.) растения. Протеомика 7: 1501–1512
PubMed CAS Google Scholar
Li K, Xu C, Li Z, Zhang K, Yang A, Zhang J (2008) Сравнительный протеомный анализ реакций фосфора в корнях кукурузы ( Zea mays L.) дикого типа и с низким содержанием фосфора. -отолерантный мутант обнаруживает характеристики корня, связанные с эффективностью фосфора. Завод J 55: 927–939
PubMed CAS Google Scholar
Li LH, Qiu XH, Li XH, Wang SP, Lian XM (2009) Профиль экспрессии генов в корнях риса при низком фосфорном стрессе.Sci China C Life Sci 52: 1055–1064
PubMed CAS Google Scholar
Лин В.Й., Лин С.И., Чиу Т.Дж. (2009) Молекулярные регуляторы фосфатного гомеостаза у растений. J Exp Bot 60: 1427–1438
PubMed CAS Google Scholar
Линдерман Р.Г., Дэвис Е.А. (2001) Сравнительный ответ отобранных подвоев и сортов виноградной лозы на инокуляцию различными микоризными грибами.Amer J Enol Viticult 52: 8–11
CAS Google Scholar
Liu J, Blaylock LA, Endre G, Cho J, Town CD, Vanden Bosch KA, Harrison MJ (2003) Профилирование транскриптов в сочетании с анализом пространственной экспрессии выявляет гены, участвующие в различных стадиях развития арбускулярного микоризного симбиоза. Растительная ячейка 15: 2106–2123
PubMed CAS Google Scholar
Liu J, Maldonado-Mendoza I, Lopez-Meyer M, Cheung F, Town CD, Harrison MJ (2007) Арбускулярный микоризный симбиоз сопровождается локальными и системными изменениями экспрессии генов и повышением устойчивости побегов к болезням. .Завод J 50: 529544
Google Scholar
McArthur DAJ, Knowles NR (1992) Устойчивые реакции картофеля на везикулярно-арбускулярные микоризные грибы при различных уровнях абиотического фосфора. Plant Physiol 100: 341–351
PubMed CAS Google Scholar
Маршнер М. (1995) Минеральное питание высших растений, 2-е изд. Академик, Лондон
Google Scholar
Marsoni M, Vannini C, Campa M, Cucchi U, Espen L, Bracale M (2005) Экстракция белка из тканей винограда с помощью двумерного электрофореза.Vitis 44: 181–186
CAS Google Scholar
Massoumou M, van Tuinen D, Chatagnier O, Arnould C, Brechenmacher L, Sanchez L, Selim S, Gianinazzi S, Gianinazzi-Pearson V (2007) Medicago truncatula генные ответы, специфичные для взаимодействий различных видов арбускулярной микоризы и роды Glomeromycota. Микориза 17: 223–234
PubMed CAS Google Scholar
Mathesius U, Keijzers G, Natera SHA, Weinman JJ et al (2001) Создание справочной карты корневого протеома для модельного бобового растения Medicago truncatula с использованием базы данных меток экспрессированной последовательности для фингерпринта пептидной массы.Протеомика 1: 1424–1440
PubMed CAS Google Scholar
Менге Д. Д., Раски Л. А., Лидер Л. В., Джонсон Н. О., Джонс Дж. Дж., Кисслер ХКЛ (1983) Взаимодействие между микоризными грибами, фумигацией почвы и ростом винограда в Калифорнии. Амер Дж. Энол Витикульт 34: 117–121
Google Scholar
Meier I (2007) Состав ядерной оболочки станции: тема и вариации.J Exp Bot 58: 27–34
PubMed CAS Google Scholar
Misson J, Raghothama KG, Jain A, Jouhet J, Block MA, Bligny R, Ortet P, Creff A, Somerville S, Rolland N, Doumas P, Nacry P, Herrerra-Estrella L, Nussaume L, Thibaud MC (2005) Полногеномный анализ транскрипции с использованием генных чипов Arabidopsis thaliana Affymetrix позволил определить реакцию растений на депривацию фосфата. Proc Natl Acad Sci USA 16: 11934–11939
Google Scholar
Николау Н., Ангелопулос К., Карагианнидис Н. (2003) Влияние стресса засухи на микоризный и немикоризный виноград Каберне Совиньон, привитый на различные подвои.Expl Agric 39: 241–252
Google Scholar
Ногалес А., Агирреолеа Дж., Мария Е.С., Кампруби А., Кальвет С. (2009) Ответ микоризной виноградной лозы на прививку Armillaria mellea: развитие болезни и полиамины. Почва для растений 317: 177–187. DOI: 10.1007 / s11104-008-9799-6
CAS Google Scholar
Palma JM, Sandalio LM, Corpas FJ, Romero-Puertas MC, McCarthy I, del Río LA (2002) Протеазы растений, деградация белков и окислительный стресс: роль пероксисом.Plant Physiol Biochem 40: 521–530
CAS Google Scholar
Poirier Y, Bucher M (2002) Транспорт фосфатов и гомеостаз у Arabidopsis. В: Somerville CR, Meyerowitz EM (eds) Книга Arabidopsis. Американское общество биологов растений, Роквилл, стр. 1–35. DOI: 10.1199 / tab.0024
Google Scholar
Посо М.Дж., Азкон-Агилар С. (2007) Выявление устойчивости, вызванной микоризой.Curr Opin Plant Biol 10: 393–398
PubMed CAS Google Scholar
Raghothama KG (1999) Приобретение фосфатов. Annu Rev Plant Physiol Plant Mol Biol 50: 665–693
PubMed CAS Google Scholar
Ramagli LS, Rodriguez LW (1985) Количественная оценка количества белка в микрограммах в буфере для образцов для электрофореза в двумерном полиакриламидном геле.Электрофорез 6: 559–563
CAS Google Scholar
Recorbet G, Dumas-Gaudot E (2008) Протеомика биотрофных взаимодействий растений и микробов: симбиозы лидируют. В: Агравал Г.К., Раквал Р. (ред.) Протеомика растений. Wiley-Intersciences, Hoboken, стр. 629–642,
. Google Scholar
Recorbet G, Valot B, Robert F, Gianinazzi-Pearson V, Dumas-Gaudot E (2010) Идентификация in planta -экспрессированных белков гриба арбускулярной микоризы при сравнении протеомов корня Medicago truncatula , колонизированных с два вида Glomus .Fungal Genet Biol 47: 608–618
PubMed CAS Google Scholar
Reynier A (2002) Manual de Viticultura, 6-е изд. Mundi Prensa, Мадрид
Google Scholar
Ruepp A, Zollner A, Maier D, Albermann K, Hani J, Mokrejs M, Tetko I, Guldener U, Mannhaupt G, Munsterkotter M, Mewes HW (2004) FunCat, схема функциональных аннотаций для систематической классификации белки из целых геномов.Nucl Acids Res 32: 5539–5545
PubMed CAS Google Scholar
Робинсон Дж., Триби М.Т., Стивенсон Р.А. (1997) Фрукты, виноградные лозы и орехи. В: Reuter DJ, Робинсон JB (ред.) Анализ растений — руководство по интерпретации. CSIRO, Мельбурн, стр. 347–389
Google Scholar
Saravanan RS, Rose JKC (2004) Критическая оценка методов экстракции образцов для усиленного протеомного анализа непокорных тканей растений.Протеомика 4: 2522–2532
PubMed CAS Google Scholar
Sarry JE, Sommerer N, Sauvage FX, Bergoin A, Rossignol M, Albagnac G, Romieu C (2004) Биохимия виноградных ягод, пересмотренная после протеомного анализа мезокарпа. Протеомика 4: 201–215
PubMed CAS Google Scholar
Schenkluhna L, Hohnjec N, Niehausa K, Schmitz U, Colditz F (2010) Дифференциальный гель-электрофорез (DIGE) для количественного мониторинга изменений протеома корня Medicago truncatula , вызванных ранним симбиозом и патогенезом.J Proteomics 73: 753–768
Google Scholar
Schubert A, Cravero MC (1985) Возникновение и инфекционность везикулярно-арбускулярных микоризных грибов на виноградниках Северо-Западной Италии. Vitis 24: 129–138
Google Scholar
Slezack S, Dumas-Gaudot E, Rosendahl S, Kjöller R, Paynot M, Negrel J, Gianinazzi S (1999) Эндопротеолитическая активность в корнях гороха, инокулированных арбускулярным микоризным грибком Glomus mosseae ic и / в отношении биозащиты.Новый Фитол 142: 517–529
Google Scholar
Smith SE, Read DJ (2008) In: Smith SE, Read DJ (eds) Микоризный симбиоз. Академик, Лондон
Google Scholar
Souza PVD, Facchin H, Dias AA (2004) Развитие подвоя виноградной лозы SO4, пораженное размером срезки и арбускулярными микоризными грибами. Cienc Rural 34: 955–957
Google Scholar
St-Arnaud M, Elsen A (2005) Взаимодействие арбускулярных микоризных грибов с почвенными патогенами и непатогенными ризосферными микроорганизмами.В: Declerck S, Strullu DG, Fortin JA (eds) Культура микоризы in vitro. Серия «Биология почвы». Springer, Berlin, pp. 217–231
. Google Scholar
St-Arnaud M, Vujanovic V (2007) Влияние арбускулярного микоризного симбиоза на болезни растений и вредителей. В: Hamel C, Plenchette C (eds) Mycorrhizae в растениеводстве: применение знаний. Haworth, Binghampton, стр. 67–122
Google Scholar
Stockinger H, Walker C, Schüßler A (2009) Glomus intraradices DAOM197198 ’, модельный гриб в исследовании арбускулярной микоризы, не является Glomus intraradices .Новый Фитол. 183: 1176–1187. DOI: 10.1111 / j.1469-8137.2009.02874
PubMed Google Scholar
Tahiri-Alaoui A, Lingua G, Avrova A, Sampò S, Fusconi A, Antoniw J, Berta G (2002) Ген кулина индуцируется в корнях томатов, формирующих арбускулярные микоризы. Can J Bot 80: 607–616
CAS Google Scholar
Takeda N, Sato S, Asamizu E, Tabata S, Parniske M (2009) Субтилазы апопластных растений поддерживают развитие арбускулярной микоризы у Lotus japonicus .Завод J 58: 766–777
PubMed CAS Google Scholar
Thelen JJ (2007) Введение в протеомику: краткий исторический обзор современных подходов. В: Самадж Дж., Телен Дж. Дж. (Ред.) Протеомика растений. Springer, Berlin, pp. 1–13
Google Scholar
Тимонен С., Смит С.Е. (2005) Эффект арбускулярного микоризного гриба Glomus intraradices на экспрессию белков цитоскелета в корнях томатов.Can J Bot 83: 176–182
CAS Google Scholar
Trouvelot A, Kough JL, Gianinazzi-Pearson V (1986) Mesure du taux de mycorhization VA d’un système radiculaire ayant une signaling fonctionnelle. В: Gianinazzi-Pearson V, Gianinazzi S (eds) Les mycorhizes: physologie et génétique. INRA, Париж, стр. 217–221
Google Scholar
Valdes-Lopez O, Hernandez G (2008) Регуляция транскрипции и передача сигналов при голодании по фосфору: как насчет бобовых? J Integrat Plant Biol 50: 1213–1222
CAS Google Scholar
Валот Б., Дьё М., Рекорбет Г., Раес М., Джанинацци С., Дюма-Годо Э. (2005) Идентификация мембранно-ассоциированных белков, регулируемых арбускулярным микоризным симбиозом.Завод Мол Биол 59: 565–580
PubMed CAS Google Scholar
Валпуэста В., Ботелла М.А. (2004) Биосинтез L-аскорбиновой кислоты в растениях: новые пути для старого антиоксиданта. Trends Plant Sci 9: 573–577
PubMed CAS Google Scholar
Vierheilig H, Coughlan A, Wyss U, Piche Y (1998) Чернила и уксус, простой метод окрашивания для арбускулярно-микоризных грибов.Appl Environ Microbiol 64: 5004–5007
PubMed CAS Google Scholar
Винсент Д., Уитли М.Д., Крамер Г.Р. (2006) Оптимизация экстракции и солюбилизации белка для созревших гроздей ягод винограда. Электрофорез 27: 1853–1865
PubMed CAS Google Scholar
Vincent D, Ergul A, Bohlman MC, Tattersall EAR, Tillett RL, Wheatley MD, Woolsey R, Joets J, Schlauch K, Schooley DA, Cushman JC, Cramer GR (2007) Протеомный анализ выявляет различия между Vitis vinifera L. .резюме. Шардоне и cv. Каберне Совиньон и их реакция на дефицит воды и соленость. J Exp Bot 58: 1873–1892
Google Scholar
Вивье М.А., Преториус И.С. (2002) Генетически модифицированные виноградные лозы для винодельческой промышленности. Trends Biotechnol 20: 472–478
PubMed CAS Google Scholar
Wang W, Bianchi L, Scali M, Liangwei L, Liu L, Bini L, Cresti M (2009) Протеомный анализ b-1,3-глюканазы в тканях ягод винограда.Acta Physiol Plant 31: 597–604
CAS Google Scholar
Waschkies C, Schropp A, Marschner H (1994) Взаимосвязь между пересаживаемой виноградной лозой и корневой колонизацией виноградной лозы (Vitis sp.) Флуоресцентными Pseudomonas и эндомикоризными грибами. Растительная почва 162: 219–227
Google Scholar
Whipps JM (2001) Перспективы и ограничения микоризы в биоконтроле корневых патогенов.Can J Bot 82: 1198–1227
Google Scholar
Wulf A, Manthey K, Doll J, Perlick AM, Linke B, Bekel T, Meyer F, Franken P, Küster H, Krajinski F (2003) Транскрипционные изменения в ответ на развитие арбускулярной микоризы у модельного растения Medicago truncatula . Mol Plant Microbe Interact 16: 306–314
PubMed CAS Google Scholar
Юань Х., Лю Д. (2008) Сигнальные компоненты, участвующие в реакции растений на фосфатное голодание.J Integr Plant Biol 50: 849–859
PubMed CAS Google Scholar
Цзэн Л. Р., Вега-Санчес М. Е., Чжу Т., Ван Г. Л. (2006) Опосредованная убиквитинированием деградация и модификация белков: новая тема во взаимодействиях растений и микробов. Cell Res 16: 413–426
PubMed CAS Google Scholar
Zrenner R, Stitt M, Sonnewald U, Boldt R (2006) Биосинтез и разложение пиримидина и пурина в растениях.Анну Рев Завод Биол 57: 805–836
PubMed CAS Google Scholar
PEM® SO4 | Самозажимные стойки с резьбой | Клинч | Клинчинговая техника | Особые технологии крепления | Продукты
PEM® SO4 | Самозажимные стойки с резьбой | Клинч | Клинчинговая техника | Специальные технологии крепления | Продукты | Bossard Global- Для использования в листах с твердостью макс.HRB 88
| Марка | Марка PEM® |
| Тип бренда | Brandtype SO4 |
| Материал | Материал Нержавеющая сталь |
Все функции
- Для использования в листах с твердостью макс.HRB 88
| Марка | Марка PEM® |
| Тип бренда | Brandtype SO4 |
| Материал | Материал Нержавеющая сталь |
| Тип материала | Тип материала AISI 400 |
| Площадь | Поверхность пассивирован |
TPST1,2 перенос SO4 (2-) из PAPS в FVIII
Посттрансляционный клеточный процессинг предшественника фактора VIII (FVIII или F8) делает возможным O-сульфатирование остатков тирозина (Pittman DD et al.1992; Michnick DA et al. 1994). Биохимические характеристики показали, что рекомбинантный человеческий FVIII при метаболической метке [35S] -сульфатом при экспрессии в клетках яичника китайского хомячка (СНО) или клетках COS-1 почек обезьяны содержит шесть потенциальных сайтов сульфатирования тирозина, то есть четыре на тяжелой цепи (при аминокислотных остатков 365, 737, 738 и 742) и два в субдомене a3 легкой цепи (остатки 1683 и 1699) (Pittman DD et al. 1992; Michnick DA et al. 1994). Присутствие шести остатков сульфата тирозина в FVIII было дополнительно подтверждено с помощью сочетания жидкостной хроматографии и масс-спектрометрии с ионизацией электрораспылением (LC / ESI-MS) рекомбинантного человеческого белка FVIII, полученного из клеток почки детеныша хомяка (BHK) (Severs JC et al. al.1999) или клетки CHO (Schmidbauer S. et al. 2015). Сайт-направленный мутагенез отдельных или множественных остатков тирозина показал, что все шесть сайтов сульфатирования необходимы для модуляции активности FVIII (Pittman DD et al. 1992; Michnick DA et al. 1994). Кроме того, обработка клеток СНО, экспрессирующих FVIII, хлоратом натрия, ингибитором АТФ-сульфурилазы, участвующей в синтезе PAPS, не влияла на секрецию FVIII, но снижала функциональную активность в 5 раз, что указывает на то, что сульфатирование не требуется для секреции FVIII. (Питтман Д.Д. и др.1992). Кроме того, мутагенез Tyr1699 в Phe (Y1699F) продемонстрировал, что сульфатирование по этому остатку необходимо для высокоаффинного взаимодействия FVIII с фактором фон Виллебранда (vWF) (Leyte A et al. 1991). В отсутствие сульфатирования тирозина на уровне 1699 в FVIII сродство к vWF снижалось в 5 раз (Leyte A et al. 1991). Исследования спектра ядерного магнитного резонанса (ЯМР) комплекса между FVIII и vWF показали значительно большие изменения химического сдвига, специфичного для остатков, когда Y1699 был сульфатирован, что дополнительно подчеркивает важность сульфатирования FVIII по Y1699 для аффинности связывания с vWF (Dagil L et al.2019). Значение сульфатирования FVIII в Y1699 in vivo становится очевидным из-за присутствия мутации Y1699F, которая вызывает умеренную гемофилию A, вероятно, из-за снижения взаимодействия с vWF и уменьшения периода полужизни в плазме (Higuchi M et al. 1990; van den Biggelaar M et al.2011). Сульфатирование по тирозиновым остаткам 365 и 1683 увеличивало активность FVIII за счет увеличения скорости расщепления тромбина в соседних сайтах расщепления тромбина 391 и 1708, соответственно (Michnick DA et al. 1994). Мутация тирозиновых остатков 737, 738 и 742 не влияла на скорость активации тромина, даже несмотря на то, что скорость расщепления на Arg759 была немного снижена (Michnick et al.1994). Способствовать. более низкая активность образования FXa (86% активности дикого типа) и более низкая активность свертывания (51% активности дикого типа) наблюдались для тройного мутанта FVIII по остаткам Tyr 737, 738 и 742 (Michnick et al. 1994). Этот результат контрастирует с другим исследованием, в котором не было обнаружено функциональных различий между полноразмерным FVIIl, не содержащим сульфатирования по одному или нескольким из этих трех остатков (Y737, Y738 и Y742), и полностью сульфатированной формой FVIII (Mikkelsen J et al. 1991).
О-сульфатирование тирозина белка является обычной посттрансляционной модификацией, которая катализируется тирозил протеин сульфотрансферазой (TPST) (Moore KL 2003; Yang YS et al. 2015). У людей были идентифицированы две изоформы TPST, названные TPST1 и TPST2 (Ouyang Yb et al. 1998; Mishiro E et al. 2006). Было показано, что фермент катализирует перенос сульфата от универсального донора сульфата аденозин-3′-фосфат-5′-фосфосульфата (PAPS) к гидроксильной группе остатка пептидилтирозина с образованием сложного эфира тирозина O4-сульфата и 3 ‘, 5’- ADP (Lee RW и Huttner WB 1983).Структурные исследования показали, что человеческие TPST обладают одним и тем же каталитическим механизмом (Teramoto T. et al. 2013; Tanaka S. et al. 2017). В клетках млекопитающих было обнаружено, что тирозин-O-сульфатирование мембранных и секреторных белков происходит в сети транс-Гольджи, а биохимические исследования показали, что фермент связан с мембраной (Lee RW & Huttner WB 1985; Baeuerle PA & Huttner WB 1987). .
| Kinkenberg et al 2018 Chem Geol.gif (значок) | [8,6 КБ] 28 августа 2018, 13:04 | Опубликовано 25 августа 2018 г. Доступно в OpenAccess с 25 августа 2020 г. |
| Kinkenberg et al 2018 Chem Geol.jpg (icon-1440) | [150.22 КБ] 28 августа 2018, 13:04 | Опубликовано 25 августа 2018 г. Доступно в OpenAccess с 25 августа 2020 г. |
| Kinkenberg et al 2018 Chem Geol.jpg (icon-180) | [23,19 КБ] 28 августа 2018, 13:04 | Опубликовано 25 августа 2018 г.Доступно в OpenAccess с 25 августа 2020 г. |
| Kinkenberg et al 2018 Chem Geol.jpg (icon-640) | [150,22 КБ] 28 августа 2018, 13:04 | Опубликовано 25 августа 2018 г. Доступно в OpenAccess с 25 августа 2020 г. |
| Kinkenberg et al 2018 Chem Geol.pdf | [7,13 МБ] 28 августа 2018, 13:02 | Опубликовано 25 августа 2018 г. Доступно в OpenAccess с 25 августа 2020 г. |
| Kinkenberg et al 2018 Chem Geol.pdf (pdfa) | [6,03 МБ] 28 августа 2018, 13:03 | Опубликовано 25 августа 2018 г.Доступно в OpenAccess с 25 августа 2020 г. |
Avaliação de catalisadores superácidos de SO4 2- / ZrO2 Destinados à reação de esterificação do óleo de algodão para produção de biodiesel.
SILVA, F. N .; http://lattes.cnpq.br/6430458195049325; СИЛЬВА, Франсиско Нильсон да.
Аннотация:
Целью данного исследования было получение двух диоксидов циркония реакцией горения (названных ZS и ZSY), оценка двух методов пропитки сульфат-ионом на поверхности синтезированного образца диоксида циркония (ZS), чтобы получить сульфатированный катализатор с максимальной возможная кислотность, то есть низкий pH.Первоначально образцы ZS и ZSY были получены реакцией горения партии из 15 г продукта с использованием н-пропоксида циркония IV и мочевины в качестве прекурсора для получения ZS и н-пропоксида циркония IV, гексагидрата нитрата иттрия и мочевины при производстве ZSY. Синтезированный образец (ZS) показал образование моноклинной основной фазы со следами тетрагональной фазы с размером кристаллитов 25,35 нм и кристалличностью 77,6%, а образец ZYS показал тетрагональную фазу как большую часть и моноклинная фаза как вторичная, с размером кристаллитов 17.79 нм и кристалличность 76,6%. Для сульфатированных образцов те же родственные фазы наблюдались также в образцах ZS и ZSY со следующими размерами кристаллитов и кристалличностью: 25,56 нм и 77,3% для образца ZS-IMA и 26,63 нм и 76,5% для образца ZSY-IMA. Метод пропитки сульфат-ионом на поверхности Отобранный образец (SZ), который показал лучший индекс pH, был IMA, равным 2,2 по сравнению с 3,6 по методу IFM (отброшено). Наилучший метод сульфатирования, использованный в образце ZS, также был использован в синтезированном образце ZSY и в коммерческом образце (ZC) моноклинной фазы, предоставленном компанией Saint-Gobain Zirpro, для целей сравнения.Оба образца имели гомогенную морфологию, состоящую из мелких частиц приблизительно сферической формы, нанометровых (с размерами частиц от 44 до 58 нм, согласно MEV, DG и BET) и более выраженного уплотнения после сульфатирования по изображениям MEV. Хроматографический анализ показал конверсию этиловых эфиров 27,57; 28,31 и 32,16% для несульфатированных образцов и вариации от 80,36 до 99,60% для сульфатированных образцов, соответственно. Результаты показали, что различие в обоих методах пропитки было существенным для выбора наилучшего метода пропитки сульфат-ионом в ZrO2, что в значительной степени способствовало выбору наилучшего метода пропитки, а вместе с тем и курсу исследования.Изменения во времени реакции сделали это можно идентифицировать катализатор (ZSY), который благоприятствовал каталитическому процессу, достигая средняя конверсия этиловых эфиров 98,57%, что на 3,11% выше целевого показателя, установленного Национальным агентством нефти, природного газа и биотоплива (Agência Nacional de Petróleo, Gás Natural e Biocombustíveis — ANP). Повторное использование имело среднее снижение содержания сложного эфира примерно на 6,05% для каждого цикла повторного использования катализатора, что можно компенсировать добавлением меньшего количества сульфат-иона в повторно используемый катализатор по сравнению с пропиткой сульфат-ионом в катализаторе повторного использования. оригинальный катализатор (без повторного использования).
| [1] | Кан ЯН, Сяоруй ШУАЙ, Хуачао ЯН, Цзяньхуа Янь, Кефа ЦЕН. Электрохимические характеристики суперконденсаторов из активированного графенового порошка с использованием ионного жидкого электролита при комнатной температуре [J]. Acta Phys. -Чим. Син., 2019, 35 (7): 755-765. |
| [2] | Цзин ЦЗОУ, Чжи-Линь ФАН, Ли-Ша ЦЗЯН, Дан-Хун ЧЖОУ.Теоретическое исследование адсорбции растворителя на активном центре Ti-Peroxo в цеолите Ti-MWW [J]. Acta Phys. -Чим. Син., 2016, 32 (4): 935-942. |
| [3] | Дун Чжэн, Сян-Ай Юань, Цзин М.А. Рационализация pH-зависимого спектра поглощения метилового красного o в водных растворах: расчет TD-DFT и экспериментальное исследование [J].Acta Phys. -Чим. Син., 2016, 32 (1): 290-300. |
| [4] | Чжао Чэн-Сяо, ВАН Хай-Тао, Ли Мин. Прогресс исследований корреляции между свойствами гелеобразования и параметрами растворителя [J]. Acta Phys. -Чим. Син., 2014, 30 (12): 2197-2209. |
| [5] | ГУ Цзя-Фан, ЧЕН Вэнь-Кай.Адсорбция иона уранила на гидроксилированной поверхности α -кварц (101) [J]. Acta Phys. -Чим. Син., 2014, 30 (10): 1810-1820. |
| [6] | Ли Цзя, Сюй Вэнь-Ли, Ху Цзин, Лин Мин, Яо Цзянь-Хуа. Механизм реакции гидролиза метаболизма 2,4-дихлорфеноксиуксусной кислоты [J].Acta Phys. -Чим. Син., 2013, 29 (09): 1923-1930. |
| [7] | НИН Хуэй, ХОУ Минь-Цян, МЭЙ Цин-Цин, ЯН Дэ-Чжун, ХАНЬ Бу-Син. Фазовое поведение систем 1-бутил-3-метилимидазолия гексафторфосфат + вода + спирт [J]. Acta Phys. -Чим. Син., 2013, 29 (04): 678-682. |
| [8] | ГУ Ли-Мэй, КУАНГ Юань-Цзян, Ян Сяо-Дань, Ю Янь-Лун, Яо Цзян-Хун, ЦАО Я-Ань.Фотокаталитическое восстановление CO 2 в CH 4 с использованием SrB 2 O 4 Катализатор [J]. Acta Phys. -Чим. Син., 2013, 29 (02): 397-402. |
| [9] | Ма Чи-Ченг, ПУ Мин, Вэй Мин, Ли Цзюнь-Нан, Ли Чжи-Хун. Два возможных пути фотореакции при оптической изомеризации L-валина [J].Acta Phys. -Чим. Син., 2012, 28 (08): 1830-1836. |
| [10] | ГУ Цзя-Фан, Лу Чун-Хай, ЧЕН Вэнь-Кай, ЧЕН Юн, Сюй Кэ, ХУАН Синь, Чжан Юн-Фань. Электронное строение карбонатных комплексов уранила (VI) в водной фазе. [J]. Acta Phys. -Чим. Син., 2012, 28 (04): 792-798. |
| [11] | Сюй Цзун-Пин, Чжао Янь-Инь, Ван Хуэй-Ган, Чжэн Сюй-Мин.Исследование структурной динамики возбужденного состояния N -метилпиррол-2-карбоксальдегида и его растворяющего действия с помощью резонансной рамановской спектроскопии и функциональной теории плотности [J]. Acta Phys. -Чим. Син., 2012, 28 (01): 65-72. |
| [12] | Ли Мин-Цзе, Лю Вэй-Ся, ПЕН Чун-Жун, Лу Вэнь-Конг. Метод первых принципов для прогнозирования окислительно-восстановительных потенциалов нуклеооснований и метаболитов в водном растворе [J].Acta Phys. -Чим. Син., 2011, 27 (03): 595-603. |
| [13] | ВАН Чжи-Цян, САН Вэнь-Цзин, Чу Вэй, Ю Лян-Цзюнь. Механизм реакции и действие растворителей при эпоксидировании стирола пероксидом водорода [J]. Acta Phys. -Чим. Син., 2011, 27 (02): 322-328. |
| [14] | ПЕН Кай-Мэй, Шао Вэнь-Ли, Ван Хуа-Хуа, ИН Сяо, Ван Хуэй, Цзи Лян-Нянь, Лю Хай-Янг.Влияние растворителей на спектральные свойства коррола галлия. [J]. Acta Phys. -Чим. Син., 2011, 27 (01): 199-206. |
| [15] | ЧЖАНЬ Чан-Го. Разработка и применение первых принципов электронного строения молекул в растворах на основе модели полностью поляризуемого континуума
[J]. |